

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The incidence of chronic kidney disease (CKD) has increased in recent years, with more people suffering from end stage renal failure [1]. A common pathologic feature of end stage kidney disease is renal tubulointerstitial fibrosis characterized by transforming growth factor β (TGF-β)/Smad signaling-mediated extracellular matrix (ECM) accumulation [23]. TGF-β/Smad signaling is a master regulatory pathway of profibrotic genes such as α-smooth muscle actin (α-SMA), plasminogen activator inhibitor 1 (PAI-1), and type 1 collagen [45]. For this reason, much effort has been placed into finding an effective strategy for inhibiting TGF-β/Smad signaling to treat renal tubulointerstitial fibrosis [67].
Thiazolidinediones (TZDs), synthetic peroxisome proliferator-activated receptor γ (PPARγ) agonists, are often used to manage type 2 diabetes mellitus (T2DM) via regulation of glucose and lipid metabolism [8]. TZDs also affect a diverse range of activities including cell proliferation, apoptosis, inflammation, and oxidative stress responses [910]. Recent studies have demonstrated beneficial effects of TZD on various renal injuries. Treatment with PPARγ agonists has protective effects against both diabetic and non-diabetic origin CKD [1112]. Although glycemic and lipid control can contribute to their protective renal effect, recent evidence suggests that upregulation of PPARγ expression in the kidney itself provides additional renal benefits by reducing TGF-β-induced ECM production, maintaining podocyte numbers and function, and regulating inflammatory cell infiltration [1314].
Lobeglitazone is a new PPARγ agonist with a TZD moiety and substituted pyrimidines, currently used to treat T2DM after completing clinical trials [15]. Phase III clinical trial data show that lobeglitazone treatment resulted in an approximately 0.6% to 0.74% decrease in glycated hemoglobin compared with that of the placebo [1617]. It is administered as a once-daily dose and mainly excreted in feces, reducing concerns of bladder cancer unlike the classic TZD pioglitazone. As another antidiabetic agent, it is necessary to evaluate additional effects of lobeglitazone on diabetic micro/macrovascular complications. A recent study has provided evidence on the cardiovascular protective role of lobeglitazone in the proliferation and migration of vascular cells. In the balloon injury rat model, lobeglitazone-treated rats showed less neointimal formation in the carotid artery than placebo-treated rats. Lobeglitazone treatment also reduced the atheromatous burden in the aorta of apolipoprotein E knockout mice fed a high-fat and high cholesterol diet [18]. However, there are no human or animal studies on any potential renal protective effects of lobeglitazone. The effects of lobeglitazone on renal tubulointerstitial fibrosis have also not been studied.
In the present study, we evaluated whether lobeglitazone had antifibrotic effects on renal tubulointerstitial disease in unilateral ureteral obstruction (UUO) mice, a model of renal tubulointerstitial fibrosis. We also examined the antifibrotic properties of lobeglitazone in vitro.
METHODS
Experimental design
C57BL6 mice were pretreated with 1 mg/kg lobeglitazone (Chung Kun Dang Pharmaceutical Corp., Seoul, Korea) by gavage daily for 3 days. For UUO-induced renal fibrosis, the left ureter of mice was doubly ligated. UUO was performed as previously described [19]. After UUO, C57BL6 mice were treated with 1 mg/kg lobeglitazone by gavage for 7 days consecutively. Seven days after UUO and lobeglitazone treatment, mice were euthanized, and their left kidneys were removed, cut in thirds, fixed in 4% paraformaldehyde, and either embedded in paraffin for histologic examination or frozen in liquid nitrogen for the isolation of protein or RNA. All procedures were performed in accordance with institutional guidelines for animal research [6].
Histologic and morphologic analysis
Histologic and morphologic analysis was performed as previously described [19]. Histochemical staining was performed with hematoxylin/eosin and sirius red. Immunohistochemical staining was performed using primary antibodies against p-Smad3 (1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), α-SMA (1:500; Sigma, St. Louis, MO, USA), PAI-1 (1:500; BD Biosciences, San Jose, CA, USA), and type 1 collagen (1:500; Abcam, Cambridge, UK), followed by horseradish peroxidase-conjugated anti-mouse or anti-rabbit immunoglobulin G secondary antibodies (Dako, Glostrup, Denmark). Quantification of renal fibrosis was measured as previously described [19].
Cell culture
NRK-49F normal rat kidney fibroblasts and rat mesangial cells (RMCs) were purchased from the American Type Culture Collection (Manassas, VA, USA). NRK-49F cells were cultured in 5% CO2/95% air at 37℃ in Dulbecco's modified Eagle's medium (DMEM; Gibco-BRL, Grand Island, NY, USA) supplemented with 5% fetal bovine serum (FBS; Hyclone, Logan, UT, USA) and antibiotics. RMCs were cultured in 5% CO2/95% air at 37℃ in DMEM. The medium was supplemented with 15% FBS and 0.4 mg/mL G418. Cells were rendered quiescent by incubation for 24 hours in medium supplemented with 0.5% FBS. Cells were treated with medium containing 0.5% FBS with or without TGF-β (5 ng/mL; R&D Systems, Minneapolis, MN, USA) for 24 hours. Cells were incubated with lobeglitazone (10 µM) for 24 hours. Cells were subsequently processed for the isolation of RNA or protein as described below.
Western blot analysis
Western blot was performed as previously described [19]. Membranes were incubated with anti-p-Smad3 (1:1,000; Cell Signaling Technology, Danvers, MA, USA), anti-Smad3 (1:1,000; Cell Signaling Technology), anti-PAI-1 (1:1,000; BD Biosciences), anti-α-SMA (1:1,000; Sigma), and anti-type I collagen (1:1,000; Abcam) polyclonal antibodies at 4℃ with gentle shaking overnight. Antibodies were detected by horseradish peroxidase-linked secondary antibody (Santa Cruz) using an Enhanced Chemiluminescence Western Blotting Detection System, in accordance with the manufacturer's instructions (Millipore, Billerica, MA, USA) [19]. The membrane was reblotted with anti-β-tubulin antibody (Applied Biological Materials Inc., Richmond, BC, Canada) to verify equal protein loading in each lane. Densitometric measurements of the bands were performed using UN-SCAN-IT digitizing software (Silk Scientific Corp., Orem, UT, USA).
Quantitative real-time reverse transcription polymerase chain reaction
Total RNA isolation and quantitative real-time reverse transcription polymerase chain reaction (RT-PCR) was performed as previously described [19]. Primers were designed using AB StepOne software version 2.1 (Applied Biosystems, Foster City, CA, USA) and were based on the relevant sequences from GenBank as follows: mouse α-SMA (GenBank accession NM_007392.3; sense, 5´-CAGGCTGTGCTGTCCCTCTA-3´; antisense, 5´-CGGCAGTAGTCACGAAGGAA-3´), mouse PAI-1 (GenBank accession NM_008871.2; sense, 5´-AATCCCACACAGCCCATCA-3´; antisense, 5´-GGACCACCTGCTGAAACACTTT-3´), mouse type 1 collagen (GenBank accession NM_ 007742.3; sense, 5´-GCCTTGGAGGAAACTTTGCTT-3´; antisense, 5´-GCACGGAAACTCCAGCTGAT-3´), mouse glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (GenBank accession NM_008084.2; sense, 5´-GAAGGGTGGAGCCAAAA G-3´; antisense, 5´-GCTGACAATCTTGAGTGAGT-3´), rat α-SMA (GenBank accession NM_031004.2; sense, 5´-GCACTACCATGTACCCAGGCAT-3´; antisense, 5´-TGCGTTCTGGAGGAGCAATAA-3´), ratPAI-1 (GenBank accession NM_012620.1; sense, 5´-CACCCCTTCCAGAGTCCCATA-3´; antisense, 5´-GCTGAAACACTTTTACTCCGAAGTT-3´), rat type 1 collagen (GenBank accession NM_053304.1; sense, 5´-GTGCGATGGCGTGCTATG-3´; antisense, 5´-TCGCCCTCCCGTTTTTG-3´), and rat GAPDH (GenBank accession NM_017008.4; sense, 5´-TGCCGCCTGGAGAAACC-3´; antisense, 5´-AGCCCAAGGATGCCCTTTAGT-3´). The housekeeping gene GAPDH was used as an internal control.
In vitro transient transfection and reporter assay
Transient transfection and reporter assay was performed as previously described [20].
RESULTS
Lobeglitazone ameliorates UUO-induced renal tubulointerstitial fibrosis
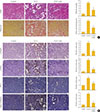
The effects of lobeglitazone on renal tubulointerstitial fibrosis were evaluated using the UUO model. As shown in Fig. 1A, hematoxylin/ eosin and sirius red staining showed that vehicle-treated UUO kidneys exhibited prominent renal tubular atrophy and tubulointerstitial fibrosis. By contrast, lobeglitazone-treated UUO kidneys showed marked attenuation of UUO-induced tubular atrophy and tubulointerstitial fibrosis (Fig. 1A).
Lobeglitazone suppresses the interstitial expression of profibrotic molecules
Given that TGF-β/Smad3 is a well-known mediator in the development of renal tubulointerstitial fibrosis, we examined the effects of lobeglitazone on the levels of Smad3 phosphorylation and Smad3 target genes including α-SMA, PAI-1, and type 1 collagen. The results showed that positively stained areas for phosphorylated Smad3, α-SMA, PAI-1, and type 1 collagen were evidently increased in the damaged tubules of UUO kidneys, but these were significantly reduced by lobeglitazone treatment (Fig. 1B).
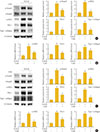
The effects of lobeglitazone on fibrotic gene expression were further confirmed by quantitative RT-PCR and Western blot analysis. Consistent with the immunohistochemical analysis, the protein abundance of PAI-1, α-SMA, and type 1 collagen was lower in the kidneys of mice administered lobeglitazone than in vehicle-treated kidneys (Fig. 2A). Moreover, UUO-induced Smad3 phosphorylation was markedly suppressed in the kidneys of lobeglitazone-treated mice (Fig. 2A). The mRNA expression levels of these genes in the kidneys of lobeglitazone-treated mice were also markedly lower than in vehicle-treated mice (Fig. 2B).
Lobeglitazone inhibits profibrotic gene expression through inhibition of TGF-β/Smad3 signaling
To examine the mechanism responsible for the antifibrotic effects of lobeglitazone, we examined whether lobeglitazone inhibits TGF-β-stimulated Smad3 signaling in cultured renal cells including NRK-49F cells and RMCs. As expected, TGF-β treatment increased mRNA and protein levels of PAI-1, α-SMA, and type 1 collagen, and induced Smad3 phosphorylation. Lobeglitazone-treated NRK-49F cells showed markedly inhibited TGF-β-stimulated profibrotic gene expression and Smad3 phosphorylation (Fig. 3A, B). Consistent with the effects in NRK-49F cells, lobeglitazone also suppressed TGF-β-stimulated Smad3 phosphorylation, PAI-1, α-SMA, and type 1 collagen in RMCs (Fig. 3C, D).
To determine whether lobeglitazone-induced suppression of profibrotic gene expression is mediated at the transcriptional level, we examined whether lobeglitazone treatment inhibits TGF-β/Smad-stimulated PAI-1 promoter activity. Indeed, lobeglitazone treatment successfully inhibited TGF-β and ALK5/Smad3, 4-stimulated PAI-1 promoter activity both in NRK-49F cells and RMCs (Fig. 4). These results indicate that lobeglitazone has an antifibrotic effect through the inhibition of TGF-β-stimulated Smad3 transcriptional activity on its target genes.
DISCUSSION
The study presented here shows that lobeglitazone treatment attenuates renal fibrosis in UUO mice. Lobeglitazone inhibited UUO-induced profibrotic gene expression including PAI-1, α-SMA, and type 1 collagen. The antifibrotic effects of lobeglitazone were associated with inhibition of the TGF-β/Smad3 signaling pathway.
PPARγ agonists are widely used as antidiabetic agents through improved insulin sensitivity and lipid metabolism [21]. On the basis of their wide range of metabolic benefits, PPARγ agonists can ameliorate diabetic nephropathy in animal and human studies [22]. Recently, several lines of evidence show that PPARγ agonists can reduce acute kidney injury, indicating that their renal protective properties may be partially independent of metabolic factors. For instance, activation of PPARγ reduces glomerulosclerosis and apoptosis via regulation of inflammation in various animal models of non-diabetic nephropathy including 5/6 nephrectomy [12], passive Heymann nephritis [23], cisplatin-induced renal damage [24], and ischemia/reperfusion injury [25]. Additionally, rosiglitazone, pioglitazone, and troglitazone have a role in reducing renal tubulointerstitial fibrosis in the UUO model [26272829]. Rosiglitazone inhibits renal tubulointerstitial fibrosis through inhibiting interstitial macrophage infiltration, downregulating the expressions of TGF-β and its down-stream target genes, and up-regulating the BMP-7 expression [26]. Several groups examined antifibrotic effects of pioglitazone and interaction with angiotensin receptor antagonists such as L158809 and candesartan in the UUO model [2728]. Pioglitazone and candesartan have additive protective effects on renal fibrosis, but the synergism between pioglitazone and L158809 is not clear [27]. Troglitazone attenuates renal interstitial fibrosis and inflammation in dose dependent manner by down regulation of TGF-β signaling pathway in the model of UUO. In accordance with these findings, our present study showed that lobeglitazone also has a protective effect on renal tubulointerstitial fibrosis and UUO-induced tubular atrophy. Furthermore, lobeglitazone inhibited the expression of well-known TGF-β target genes, such as PAI-1, α-SMA, and type 1 collagen, as well as its major effector, phosphorylated Smad3, in the kidneys of UUO mice.
The TGF-β/Smad signaling pathway is a primary pathogenic factor that drives glomerular and tubulointerstitial fibrosis in the kidney by stimulating the synthesis of ECM molecules and by decreasing ECM degradation [4]. Although mesangial cells express Smad1, 2, 3, 4, and 7, accumulating evidence demonstrates that Smad3 is mainly implicated in a pathogenic role in TGF-β-mediated renal fibrosis [303132]. Recent studies show that several PPARγ agonists inhibit TGF-β-stimulated ECM production, indicating that PPARγ agonists attenuate renal tubulointerstitial fibrosis by inhibiting the TGF-β/Smad3 signaling pathway [33]. Pioglitazone inhibits renal tubulointerstitial fibrosis and infiltration of interstitial macrophages by regulating transcription of PAI-1in UUO mice [28]. Rosiglitazone treatment inhibits inflammatory reactions and renal fibrosis by reducing the overexpression of endogenous endothelin-1, cyclooxygenase-2, and TGF-β in deoxycorticosterone acetate-salt hypertensive rats [34]. Our study adds evidence that another novel PPARγ agonist, lobeglitazone suppresses tubulointerstitial fibrosis after UUO in mice by inhibiting TGF-β/Smad3 signaling.
In conclusion, this study demonstrates that lobeglitazone has a renoprotective effect on UUO-induced renal fibrosis through inhibition of the TGF-β/Smad3 pathway. Our results suggest that lobeglitazone could play a therapeutic role in CKD, providing rationale for further clinical trials to evaluate the efficacy of lobeglitazone in the treatment of CKD including renal tubulointerstitial fibrosis.


 XML Download
XML Download