

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
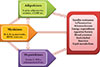
Obesity significantly increases the risk of cardiometabolic diseases, such as type 2 diabetes, dyslipidemia, hypertension, coronary heart disease, and stroke. Furthermore, recent evidence has shown that obesity is a risk factor for nonalcoholic fatty liver disease (NAFLD), obstructive sleep apnea, dementia, and several types of cancers [1]. The main known function of adipose tissue is to store excess calories in the form of triglycerides and to release them during fasting and cold exposure. Moreover, adipose tissue has been established to be an endocrine organ that secretes adipokines, which are bioactive peptides that control systemic metabolism and energy homeostasis [2]. Organokines, which are predominantly produced and se-creted by their respective tissues, affect whole body metabolism through autocrine, paracrine, and endocrine activities (Fig. 1). Autologous to adipokines, myokines from skeletal muscle and hepatokines from the liver also regulate a variety of biological processes and communicate with distant target organs.
ADIPOKINES
In obese individuals, the hypertrophy of adipocytes predisposes to immune cell infiltration, which produces proinflammatory cytokines, including interleukin 6 (IL-6), IL-8, and monocyte chemoattractant protein-1 (MCP-1). With increasing body weight, individuals develop adipocyte hypertrophy, hypoxia, and ectopic fat deposition, which induces the altered production and secretion of adipokines in adipose tissue [3]. Adipose tissue dysfunction and disturbed adipokine secretion may connect obesity with its metabolic, inflammatory, and cardiovascular complications [45]. Adipokines such as leptin, adiponectin, tumor necrosis factor α (TNF-α), retinol binding protein 4 (RBP4), adipocyte fatty acid binding protein (A-FABP), resistin, vaspin, apelin, chemerin, omentin, and C1q/TNF-related proteins (CTRPs) are involved in a spectrum of obesity-associated disorders. In fact, several adipokines, such as leptin, RBP4, IL-6, chemerin, and progranulin, are differentially expressed according to fat deposition [1].
Leptin is a prototype adipokine discovered in 1994 [6]. Leptin has a pivotal role in the regulation of appetite, satiety, food intake, energy expenditure, reproductive function, and fertility [7]. In the hypothalamus, leptin decreases orexigenic- and increases anorexigenic-neuropeptide synthesis, resulting in reduced appetite and body weight in animal experiments [8]. However, circulating leptin levels are proportional to body fat mass, and treatment with recombinant leptin shows only small effects on weight loss in humans [9]. These results support the concept of leptin resistance or tolerance in obese individuals. Adiponectin is a 30-kDa protein which signals via at least two adiponectin receptors, AdipoR1 and AdipoR2 [10]. Post-translational modification of adiponectin results in 180-kDa hexamers (low molecular weight) and 18- to 36-mers (high molecular weight) [1011]. Adiponectin improves insulin sensitivity and has anti-diabetic, anti-inflammatory, and anti-atherogenic properties [1]. Circulating adiponectin levels are inversely associated with components of the metabolic syndrome, including body weight, blood pressure, lipids, and insulin resistance. Furthermore, adiponectin levels show a negative relationship with visceral fat, type 2 diabetes, and cardiovascular disease (CVD) [11]. We reported that circulating adiponectin levels showed a significant negative correlation with the mean target-to-background ratio, which reflects vascular inflammation using 18F-fluorodeoxyglucose positron emission tomography (18FDG-PET) [12]. A-FABP is a novel adipokine that plays a key role in linking obesity with various features of metabolic syndrome, type 2 diabetes, and CVD [1314]. In macrophages, A-FABP potentiates toxic lipid-induced inflammation and endoplasmic reticulum (ER) stress, which may be involved in the pathogenesis of CVD [15]. Several prospective studies have shown that high A-FBAP concentrations at baseline are predictive of the risk for metabolic and vascular morbidity, as well as mortality [16]. In our previous study, A-FABP levels predicted the development of metabolic syndrome independent of traditional risk factors in Korean children [17]. Recent studies have shown that the CTRP family of proteins, paralogs of adiponectin, play a crucial role in the regulation of systemic metabolism and innate immunity [1819]. CTRP-1 to CTRP-15 show remarkable similarities in their structural and biochemical properties compared to adiponectin [18]. We observed that CTRP-3 concentrations are elevated in patients with dysregulation of glucose metabolism and are associated with various metabolic risk factors [20]. Moreover, a 3-month combined exercise program significantly affected CTPR-3 and CTRP-5 levels in obese Korean women [21]. Patients with acute coronary syndrome and stable angina pectoris had significantly lower circulating CTRP-3 levels compared to control subjects [22]. Recently, we demonstrated that CTRP9 attenuates hepatic steatosis through the alleviation of ER stress via the AMP-activated protein kinase (AMPK)-mediated induction of autophagy [23].
MYOKINES
Aging and physical inactivity result in a progressive loss of muscle mass and strength, known as sarcopenia. We have previously studied the prevalence of sarcopenia and its impact on metabolic disorders, including type 2 diabetes and NAFLD [242526]. Regular exercise is an effective way to prevent and treat chronic metabolic disorders such as type 2 diabetes and metabolic syndrome. Myokines, proteins that are released from skeletal muscle, may mediate the beneficial effects of regular physical activity [27]. In fact, exercise affects the expression in skeletal muscle and the circulating levels of a number of myokines, such as IL-6, IL-15, angiopoietin-like 4, myostatin, and irisin [28].
IL-6 is the first myokine that has been known to be a proinflammatory cytokine. Dr. Pedersen's group first demonstrated that IL-6 mRNA expression in muscle, as well as circulating levels of IL-6, were markedly increased after exercise [29]. During acute exercise, IL-6 improves insulin sensitivity by blocking proinflammatory signaling pathways in the muscle [30]. However, chronically elevated IL-6 induces insulin resistance in adipose tissue and the liver [27]. Irisin is an exercise-induced novel myokine that promotes the browning of white adipose tissue and may mediate the health-promoting effects of exercise, including improvement in glycemic control [31]. However, controversy has arisen regarding inconsistencies in the regulation of irisin after exercise [32]. We showed that circulating irisin concentrations were not different in individuals with sarcopenia and those with brown adipose tissue detectable by 18FDG-PET [33].
HEPATOKINES
The liver is a major regulator of systemic metabolism and whole body energy homeostasis. Liver-derived proteins known as hepatokines have recently emerged as novel hormones that have ambivalent roles, either aggravating insulin resistance or improving metabolic variables in the metabolic syndrome [34]. Fetuin-A is the first-known hepatokine that integrates crosstalk between the liver and target organs. Fetuin-A, a natural inhibitor of the insulin-stimulated insulin receptor tyrosine kinase, induces insulin resistance in rodents [35]. Fetuin-A was identified as an endogenous ligand for Toll-like receptor 4 through which saturated fatty acids induce proinflammatory signaling and insulin resistance [36]. Circulating fetuin-A levels are increased in obesity, metabolic syndrome, and type 2 diabetes, and are correlated with hepatic steatosis in humans [37]. Furthermore, prospective studies have shown that fetuin-A levels predict the increased risk of type 2 diabetes as well as myocardial infarction and stroke [3839]. We reported that adiponectin and salsalate improve hepatic steatosis by the inhibition of fetuin-A through the AMPK-nuclear factor-κB pathway [40]. Furthermore, we found that caloric restriction significantly decreased hepatic fetuin-A expression and its circulating levels in obese rats and humans with type 2 diabetes [41]. Fibroblast growth factor 21 (FGF21) is a central metabolic regulator that has favorable effects on glucose and lipid metabolism [42]. FGF21 is upregulated by the nuclear receptor peroxisome proliferator-activated receptor α during starvation [43]. Administration of FGF21 has been shown to decrease body weight, blood glucose, and lipid levels, as well as improves insulin resistance in animal experiments [44]. FGF21 decreases blood glucose levels by increasing glucose transporter 1 expression in human primary adipocytes and reduces triglyceride levels by inhibiting lipolysis and augmenting β-oxidation in the liver and adipose tissue [4546]. Circulating concentrations and the hepatic expression of FGF21 are increased in patients with NAFLD [47]. In a recent randomized clinical trial, treatment with LY2405319, an analog of FGF21, resulted in significant improvements in the dyslipidemia of obese human subjects with type 2 diabetes [48]. Selenoprotein P is positively correlated with insulin resistance and is upregulated in patients with type 2 diabetes [49]. In our previous study, selenoprotein P levels were elevated in patients with dysregulated glucose metabolism and were significantly associated with insulin resistance, inflammation, and carotid intima-media thickness [50]. We also reported that AMPK activators alleviate carrageenan-induced insulin resistance through the AMPK-mediated amelioration of ER stress in hepatocytes [51]. Circulating selenoprotein P levels are negatively correlated with adiponectin levels in patients with type 2 diabetes, suggesting the existence of crosstalk between a hepatokine (selenoprotein P) and an adipokine (adiponectin) [52].
CONCLUSIONS
Adipose tissue is now established as an active endocrine organ that secretes adipokines. Altered adipokine production and secretion may provide a link between adipose tissue dysfunction and obesity-related disorders. Adipokines are pivotal regulators in whole body metabolism because they are involved in impaired insulin sensitivity or secretion, inflammation, fat distribution, appetite, and satiety, as well as endothelial dysfunction and atherosclerosis. Analogous to the role of adipokines, myokines and hepatokines have also been proven to have crucial pathogenic roles in metabolic syndrome, NAFLD, type 2 diabetes, and CVD. The identification and functional characterization of novel organokines may provide important insights that could lead to novel treatments for cardiometabolic diseases.
 XML Download
XML Download