

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In mammals, the endocrine pancreas comprises a majority of the islets of Langerhans that are dispersed within the pancreas [1]. There are approximately 1,000 islets in a mouse pancreas and 1 million islets in a human pancreas [2,3]. Each islet is a round or ovoid mass, 50 to 600 µm in diameter and comprising ~50 to 3,000 cells [1,3]. Pancreatic islets are composed of four different endocrine cell types. Highly specialized cells called β-cells represent about 60% of the cells in human islets. The remaining cells are α, δ, polypeptide-producing, and epsilon cells, which produce glucagon, somatostatin, pancreatic polypeptide, and ghrelin, respectively [1,2].
The endocrine pancreas alone is responsible for insulin secretion in mammals and plays a prominent role in controlling circulating glucose levels and glucose metabolism [2,4]. Insulin is produced by β-cells in the pancreas [2,5,6,7]. Pancreatic β-cells produce, store, and finally secrete insulin in response to increased glycemia [1]. However, the modulation of insulin secretion as a function of changing metabolic demands and environmental conditions, especially levels of circulating glucose, is not completely performed by β-cells alone. It is a highly complex process, and many factors affect β-cells through vascular, paracrine, autocrine, neural, and cellular pathways [8]. Within pancreatic islets, each cell communicates in a variety of ways, including through indirect and direct mechanisms [1]. Indirect cell communication does not require cell contact but is instead mediated by an extracellular flux of molecules that simultaneously signal to multiple cells. In addition, various modalities of direct cell-to-cell communication operate within pancreatic islets. Therefore, the segregation of islets into single cells leads to a rapid loss of insulin regulation. This regulation can be restored by reestablishing cell contacts [1,9,10,11,12]. Because the integrated secretory responses of islet cells are essential for the maintenance of normal patterns of insulin secretion, several recent studies have focused on interactions between different islet cell types, especially α- and β-cells [13].
Research on pancreatic β-cell function and regulation in many cases still relies on clonal cell line monolayers, which have limited ability to mimic endogenous pancreatic islets. Using pancreatic islets themselves is the best approach. However, islet isolation is complex and time consuming. The number of islets that can be obtained by that process largely depends on the experience of the experimenters. In addition, isolated islets cannot survive longer than 2 weeks or be stockpiled. Therefore, we sought to determine whether it would be viable to use co-cultured α and β clonal cell lines instead of islets. We assessed their morphology and function and compared them to single cell cultures and pancreatic islets.
METHODS
Cell lines
Mouse α TC-6 cells and β TC-1 cells were obtained from the American Type Culture Collection (Manassas, VA, USA). Cells were grown at 37.0℃ in a 5% carbon dioxide incubator in Dulbecco's Modified Eagle's Medium (DMEM) containing 4 mM L-glutamine, 5 mM glucose, 44 mM sodium bicarbonate, 10% heat-inactivated fetal bovine serum (FBS), 15 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), and 0.1 mM nonessential amino acids.
α TC-6 cell and β TC-1 cell co-culture system
Cells were cultured in culture plates in Dulbecco's Minimal Essential Medium containing 5 mM glucose and supplemented with 44 mM sodium bicarbonate, 10% FBS, and 15 mM HEPES. Initially the cells were mixed at various α:β ratios: 1:4, 1:2, and 1:1. However, we found it difficult to observe the α-cells at ratios of 1:4 and 1:2.
Therefore, cells were mixed at a 1:1 ratio (5×105) in 6-well plates. The plates were shaken on a gyratory shaker at 37℃ (Fig. 1).
Rat islet isolation
Hank's balanced salt solution (HBSS) was used for perfusion and preservation of pancreas. Pancreatic islets were isolated using Lacy and Kostianovsky's method with modifications. Pancreases were distended by intraductal injection of cold collagenase solution (0.8 mg/mL in HBSS) into the common bile duct after occlusion of the distal end, close to the duodenum. Digestion was performed in a water bath at 37℃ for 14 minutes 20 seconds and was stopped by the addition of cold HBSS. Each pancreas was broken up by vortexing for 15 seconds. After centrifugation for 10 seconds at 1,500 rpm, the supernatant was removed. The pancreatic tissue was resuspended in 30 mL HBSS and passed through a 400-µm mesh. The filtered tissue was washed with HBSS and centrifuged on a Bicoll density gradient (1.085, 1.069, and 1.037 g/mL). The islets were cultured with M199 medium containing 10% FBS in a noncoated plate.
Morphological examination and immunostaining
The aggregates were observed at different time points with a phase-contrast microscope. α TC-6 and β TC-1 cells were plated onto cover slides. After incubation under appropriate conditions, the cells were fixed and permeabilized with Cytofix/Cytoperm (BD, San Diego, CA, USA) for 15 minutes and washed with perm/wash buffer (0.1% saponin, 1 mM CaCl2·2H2O, 1 mM MgSO4, 0.05% NaNa3, 10 mM HEPES, 1% bovine serum albumin). Cells were incubated with an anti-insulin or antiglucagon antibody overnight at 4℃ and washed with phosphate-buffered saline (PBS). Then the cells were incubated with secondary antibody (antirabbit immunoglobulin G [IgG] Alexa Fluor 488 conjugate or Alexa Fluor 555 conjugate [Cell signaling, Danvers, MA, USA]) for 1 hour at room temperature and washed with PBS. We used anti-insulin and antiglucagon antibodies isolated from rabbits instead of rats because we used mouse α TC-6 and β TC-1 cells. However, the antirabbit IgG secondary antibody stained both insulin and glucagon. Therefore, we could not use confocal microscopy to distinguish between insulin and glucagon, and we stained each of them separately. Fluorescent in situ hybridization analysis was performed using a BX-51 green fluorescence microscope (Olympus, Tokyo, Japan). Then we calculated the cellular composition of the co-culture aggregates at 24, 48, and 72 hours. The results are presented as cell number/area (%).
Quantification of secreted insulin protein by enzyme-linked immunosorbent assay
After incubation, cells were washed with PBS. To measure glucose-stimulated insulin secretion (GSIS), we starved cells for 5 hours in DMEM containing 5 mM glucose and 2% FBS. Then the medium was replaced with KRBB solution (4.74 mM KCl, 1.19 mM KH2PO4, 1.19 mM MgCl2·6H2O, 35 mM NaHCO3, 10 mM HEPES) containing 5 mM glucose or 25 mM glucose and the cells were incubated for 1 hour. To separate total insulin protein, we washed cells with PBS and lysed them with mammalian tissue lysis/extraction reagent containing a protease inhibitor. Insulin protein in the medium was measured using a Rat/Mouse Insulin ELISA Kit (Linco Research, St. Charles, MO, USA).
RESULTS
Formation of aggregates in the co-culture system and morphologic examination
Cells were mixed at a 1:1 ratio (5×105) in 6-well plates and the plates were sealed and shaken on a gyratory shaker to form aggregates. After culture, we observed the morphology with an optical microscope (×200) at 2 hours (Fig. 2A) and 48 hours (Fig. 2B). Cells were scattered at 2 hours (Fig. 2A), but numerous aggregates of heterogeneous size had formed from α TC-6 and β TC-1 cells at 48 hours (Fig. 2B). They became condensed and more strongly compacted at 72 hours. They survived well in the co-culture system, and good cell-to-cell contacts were formed using the gyratory shaker.
Immunohistochemical staining of co-cultured cells
Immunohistochemical staining for insulin and glucagon at 24 hours (Fig. 3A), 48 hours (Fig. 3B), and 72 hours (Fig. 3C) is shown in Fig. 3. β TC-1 cells showed both β-β and β-α cell contacts, whereas α TC-6 cells showed both α-α and β-α cell contacts. α TC-6 cells became compact and more spherical by 72 hours, but β TC-1 cells did not show a specific pattern in the co-culture system.
We also analyzed the cellular composition of the aggregates (Fig. 4). Of cells mixed at a 1:1 ratio (5×105), ~45% showed α-α cell contacts and ~25% showed β-α cell contacts. The percentage of β-α cell contacts was similar through 72 hours, although the total cell population increased.
Glucose-stimulated insulin secretion
GSIS from β TC-1 cells and co-cultured cells is shown in Fig. 5. Insulin secretion was increased by glucose stimulation in both cultures. The degree of GSIS was higher in β-cells cultured alone than in the α-/β-cell co-culture.
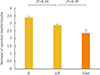
We also compared the ratio of GSIS to insulin contents in β TC-1 cells, co-cultured cells, and islets. As shown in Fig. 6, the increase in secreted insulin/insulin content was significantly lower for co-cultured cells than for β-cells only (P=0.04). Compared to islets, the α-/β-cell co-culture showed a higher ratio of GSIS to insulin content, but the difference was not statistically significant (P=0.09).
DISCUSSION
The present study demonstrated that α TC-6 cells and β TC-1 cells formed aggregates that looked like pseudoislets and survived well in our co-culture system. They showed similar GSIS to pancreatic islets. Although our morphologic study did not show a particular topography similar to the three-dimensional architecture of native islets, the co-cultured cells formed multiple aggregates with cell contacts and showed similar GSIS to islets isolated from rats.
In previous studies, different cell types showed a particular intraislet topography: β-cells were located in the central core, whereas non-β-cells surrounded the β-cells to form a mantle layer [8,13].
Most researchers study pancreatic β-cells with clonal cell line monolayers. However, the normal pattern of insulin secretion is facilitated by the maintenance of islet architecture, and the integrated secretory response of islet cells is essential for the normal pattern [14]. There have been many studies of interactions between and within pancreatic islets. It is now known that intraislet cell communication is important for adequate release of insulin and is largely achieved through gap junction-medicated communication [15]. Although the exact mechanism is unknown, several mechanisms depend on different families of integral membrane proteins, which ensure direct (cadherins, neural cell adhesion molecule, occludin, and claudins) and paracrine (pannexins) communication between β-cells and between these cells and the other islet cell types [1]. Also, other proteins (integrins) provide communication between the different islet cell types and the materials that form the islet basal laminae and extracellular matrix. The dissociation of islet cells is associated with a dramatic change in insulin secretion in response to glucose. The formation of cellular aggregates called pseudoislets marks a significant improvement in insulin secretion [1,2,8]. In the early work of Timble and others, the cellular glucagon content of dorsal islets was associated with insulin secretion [16]. Another study showed that an increase in the percentage of glucagon-secreting cells to 50% had a detrimental effect on insulin release from neighboring β-cells [17]. These findings suggest that the interaction of insulin-secreting cells and α-cells and changes in their relative proportions have marked effects on insulin release. Pseudoislets comprising insulin-secreting cell lines and α-cell lines could be a useful model for examining islet interactions and insulin secretion.
Most pseudoislet research has been conducted on clonal β-cell lines only and has isolated β-cell lines without other islet cells such as α-cells. There have been few studies of heterotypic pseudoislets containing α- and β-cell lines [8,13,18]. One study used a model containing low concentrations of α TC1 and MIN6 cells. α TC1 cells had no effect on insulin secretion from MIN6 cells in that study [18]. However, in another study, co-cultured pancreatic α- and β-cell lines exhibited contact-dependent inhibition of insulin secretion [13]. In that study, insulin-secreting β TC cells and glucagon-secreting α TC cells formed pseudoislets. Our present study showed comparable results: the co-cultured cells formed pseudoislet-like aggregates and showed decreased GSIS compared to β-cells cultured alone. It has not been clearly explained how insulin secretion from β-cells is inhibited via contact with α-cells. Previous studies have suggested several hypotheses. An increase in the α TC/β TC cell ratio in aggregate cultures was accompanied by a decrease in secreted insulin and an increase in the intracellular insulin content of the β TC component [13]. Because the intracellular insulin content of β TC cells was not affected by α TC cells in a Transwell (Cat. No. 3413, Costar, Cambridge, MA, USA) co-culture system in which direct cell-to-cell contacts were prevented by a semipermeable membrane that permitted chemical communication via secreted metabolites, the authors guessed that insulin secretion from β TC cells may possibly be inhibited as a result of contact with α TC cells. However, the authors did not completely rule out the possible effects of soluble factors. The possibility of apoptosis of β TC cells due to cell-to-cell contacts has been suggested [13]. We think that further investigation is needed to determine the precise mechanism.
In conclusion, we have shown both β-β and β-α cell contact in the co-culture system and demonstrated that co-cultured cells have similar GSIS to pancreatic islets. Although the use of isolated islets is the best way to study pancreatic function, this method is problematic for many reasons. We think that our pseudoislets, created from α- and β-cell lines using a simple method, may mimic pancreatic islets, reducing the limitations of in vitro experiments.




 XML Download
XML Download