

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
A number of genetic alterations have been shown to play a role in the development of follicular cell-derived thyroid cancer. These point mutations and translocations occur in genes of several important signaling pathways, particularly that of the mitogen-activated protein kinase (MAPK) pathway. The MAPK signaling pathway is a master regulator of numerous cellular processes including division, proliferation, differentiation, adhesion, migration, and apoptosis. B-Raf proto-oncogene (BRAF) mutations, RET/papillary thyroid cancer (PTC) rearrangements, and RAS mutations are the most common activators of the MAPK signaling pathway, with significant implications for thyroid tumorigenesis.
BRAF mutations are the most common mutations observed in PTCs, followed by RET/PTC rearrangements and RAS mutations, while follicular thyroid cancers (FTC) are more likely to harbor RAS mutations or PAX8/peroxisome proliferator-activated receptor γ (PPARγ) rearrangements. While all four of these mutations are common worldwide, the prevalence of each mutation type in thyroid cancer varies significantly, particularly between Asian and Western countries, with the prevalence of PTC significantly higher in Asian countries.
Beyond these more common mutations, alterations in the telomerase reverse transcriptase (TERT) promoter may be predictive of clinicopathologic features, as well as disease prognosis and tumorigenesis in thyroid cancer. Like other common thyroid cancer mutations, the frequency of TERT promoter mutations also appear to differ among countries, though the significance of this observation remains limited due to the small number of studies on this mutation having been conducted to date.
In this article, we review the prevalence of each of the most common mutations in Asian and Western countries, and identify the characteristics of well-differentiated thyroid cancer (DTC) in Asians.
BRAF MUTATION
BRAF, located in chromosome 7, is the most commonly mutated gene in thyroid cancers, resulting in potent activation of the MAPK pathway. The most common mutational hotspot in BRAF is T1799A in exon 15, conferring a glutamate to valine substitution at amino acid 600 (V600E) in the BRAF protein. BRAF V600E is the most common genetic alteration in PTC, exhibiting high prevalence in classic PTC and the tall cell variant, although it is generally rare in the follicular variant. Because BRAF mutations can be detected preoperatively in fine needle aspiration biopsy (FNAB) specimens, it is often used in the diagnosis of PTC, and may inform initial treatment strategies. Furthermore, this mutation has emerged as a promising prognostic factor for PTC [12], although the prognostic value of this mutation is still inconclusive [34].
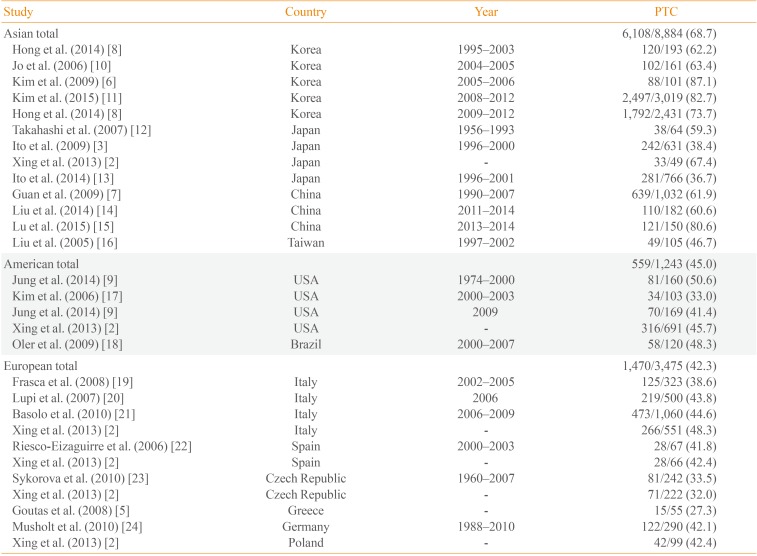
The overall prevalence of BRAF mutations is ~45% (range, 27.3% to 87.1%) [56], with prevalence significantly higher in Asia, especially Korea, relative to Western countries (Table 1) [2356789101112131415161718192021222324]. Although the mechanisms underlying this difference in BRAF mutation frequencies are not well understood, a recent theory suggests that these differences may be associated with higher iodine intake in the Asian populations. Average iodine intakes were 138 to 353 µg/day in the United States [25], 45.3 µg/day in Germany [26], and 226 and 163 µg/day for women and men, respectively, in the United Kingdom [27]. Meanwhile, Japanese and Korean iodine intakes far exceed that of most other countries: 1,565 µg/day in Japan [28] and 479 µg/day in Korea [29]. Furthermore, high iodine intake has been shown to be significantly associated with the occurrence of BRAF mutation [7], though exceptions do exist, including lower BRAF mutation rates in Japan relative to Korea. One possible explanation for this discrepancy may be that of chronic thyroiditis, which is more prevalent in the Korean population. Incidence of Hashimoto's thyroiditis is strongly correlated with the development of PTC [30]. Because the prevalence of Hashimoto's thyroiditis is high in Korea, this positive correlation may provide an explanation for the high incidence of PTC in this country. However, as Hashimoto's thyroiditis is associated with genetic alterations other than BRAF mutations, such as rearrangements of RAS, ERK, and RET/PTC [31], the relationship between BRAF mutations in PTC and chronic thyroiditis requires further assessment.
While geographic differences in the incidence of BRAF mutations are well established, the prevalence of these mutations has changed over time. A recent publication from our laboratory revealed an increase in BRAF-associated thyroid cancers from 62.2% to 73.7% over the last two decades in Korea [8]. Similarly, in the United States, the overall prevalence of BRAF mutations remained stable for an extended period of time (~46%) but increased sharply from 50.0% to 76.9% in the classic papillary form of PTC over the last four decades [9]. More studies on the changes in the mutational rates and its clinical significance will be needed.
RAS MUTATION
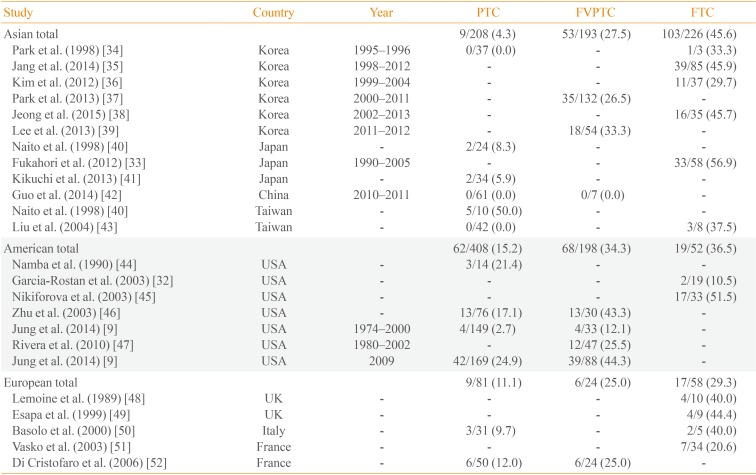
RAS mutations are the second most common genetic alteration in thyroid cancer. The RAS gene encodes a family of three isoforms: NRAS, HRAS, and KRAS. Thyroid neoplasms have been associated with mutations in all three isoforms of the RAS gene, although most studies have reported a predominance of NRAS61. RAS point mutations are commonly observed in FTC, as well as the follicular variant PTC. The frequency of RAS mutations in FTC ranges from 10.5% to 56.9% [3233], and is slightly more common in Asia (45.6%) than in Western countries (36.5% in the Americas, and 29.3% in Europe). In contrast, the frequency of RAS mutations in PTC is much lower in Asia (Table 2) [9323334353637383940414243444546474849505152]. This low frequency of RAS mutations has remained relatively stable over time, which is likely to be due to the lower prevalence of follicular variant PTC in this population. In contrast, a study from the United States reported an increase in the proportion of RAS mutation-positive from 2.7% between 1974 and 2000 to 24.9% in 2009, due in part to an increase in the percentage of patients presenting with the follicular variant histology [9].
RAS mutations have been reported in the full spectrum of thyroid neoplasms, limiting the clinical diagnostic value of these mutations. Because it is difficult to differentiate specific types of follicular lesions in thyroid FNAB samples, the diagnostic use of RAS mutations in FNAB specimens remains controversial. The prognostic value of RAS mutations is also unclear, although some evidence suggests that RAS-positive thyroid cancers may be at risk for tumor dedifferentiation, a less favorable prognosis, and metastatic behavior, particularly with regard to bone metastasis [3233].
RET/PTC REARRANGEMENT
Rearrangements of the RET proto-oncogene are commonly seen in PTC, and have been shown to play a role in disease pathogenesis. To date, 13 different types of RET/PTC rearrangements have been reported, though RET/PTC1 and RET/PTC3 account for more than 90% of all rearrangements. The relationship between radiation exposure and RET/PTC rearrangement has been established [5354], with RET/PTC rearrangements frequently observed in PTC patients who have received significant doses of external radiation, such as those affected by the Chernobyl nuclear accident. Elevated levels of childhood PTC are well documented in post-Chernobyl contaminated areas, accompanied by a high prevalence of RET/PTC rearrangements. Rapid proliferation of thyroid cells may account for the high sensitivity to radiation-induced RET/PTC rearrangements among children, although RET/PTC rearrangements also occur more frequently in children and young adults not exposed to radiation [55].
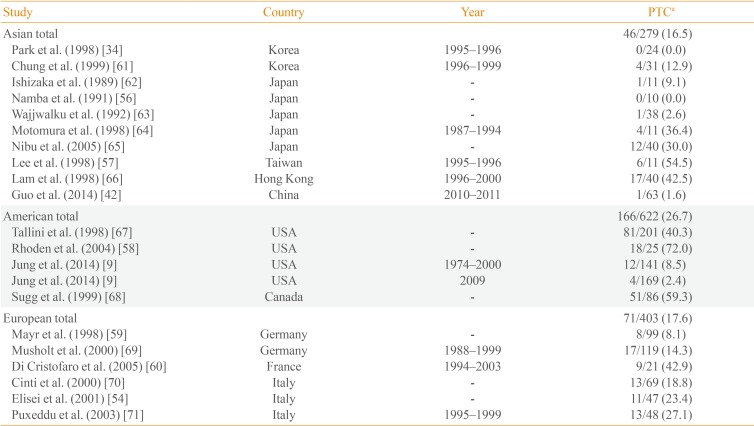
The prevalence of RET/PTC rearrangements in PTC varies widely in different populations (range, 0% to 86.8% [345356]), with significant variability in mutational frequency even within the same geographical regions (0% to 54.5% in Asia [345657], 2.4% to 72.0% in the Americas [958], and 8.1% to 42.9% in Europe [5960]). These discrepancies may be due to the small size of the studies; when this variability is taken into account, the prevalence of RET/PTC rearrangements in Asia is generally low (16.5%) (Table 3) [934425456575859606162636465666768697071].
This wide range of the prevalence rates seen in these studies may reflect not only the geographic variability but also the effect of different detection methods. A variety of methods have been used to identify RET/PTC rearrangements, including reverse transcription polymerase chain reaction methods, Southern blot analysis, and fluorescence in situ hybridization. Zhu et al. [72] demonstrated that different detection methods could result in significant variability in the detection of RET/PTC rearrangement.
PAX8/PPARγ REARRANGEMENT
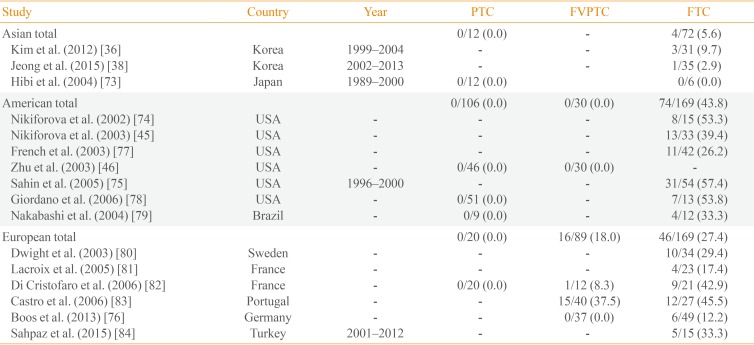
PAX8/PPARγ rearrangements occur as a result of an intrachromosomal translocation between most of the coding sequence of PAX8 (2q13) and the entire coding exons of PPARγ1 (3p25). The fusion gene appears to be an oncogene, and results in production of a PAX8/PPARγ fusion protein (PPFP). The PAX8/PPARγ fusion gene is most commonly found in FTC, the follicular variant PTC, and benign follicular adenomas, though the prevalence of these rearrangements varies significantly among studies. The mean frequency in FTC is 5.6% in Asia, 43.8% in the Americas, and 27.4% in Europe (Table 4) [36384546737475767778798081828384]. The low frequency of PAX8/PPARγ rearrangements in Asia is particularly noteworthy, with one Japanese study failing to identify a single PAX8/PPARγ rearrangement in FTC [73].
No evidence exists linking PAX8/PPARγ rearrangements with clinical outcomes in FTC. Multiple studies have reported no correlation between PAX8/PPARγ rearrangements and clinical variables such as gender, age, tumor size, lymph node metastasis, recurrence, or mortality [747576].
Despite the lack of clinical associations, PPARγ remains an attractive therapeutic target in thyroid cancer. Although PPARγ agonists have shown promising results in both in vitro and in vivo studies [8586], the results of these studies have been inconclusive. Larger studies with long-term follow-up will be needed to clarify the efficacy and availability of PPARγ agonists in PPFP thyroid cancer.
TERT PROMOTER MUTATION
Somatic mutations in the TERT promoter have been identified in many human malignancies including thyroid cancer. Mutations in the TERT promoter have been shown to increase telomerase activity, which protects the telomere repeats from erosion and plays a key role in cellular immortality and tumorigenesis [87]. TERT promoter mutations were mainly found in two hotspots, located -124 (chr5: 1,295,228C>T) and -146 bp (chr5: 1,295,250C>T) upstream of the gene transcription starting site. These mutations were recently shown to be more prevalent in aggressive thyroid cancers, and were associated with poor prognosis as well as high-risk clinicopathologic features [888990]. Therefore, TERT promoter mutation has received considerable attention as a novel prognostic biomarker. TERT promoter mutations have been shown to coexist with other tumorigenic alterations, such as BRAF or RAS mutations. Indeed, the coexistence of BRAF mutations and TERT promoter mutations has been identified as an indicator of the worst prognosis [8891].
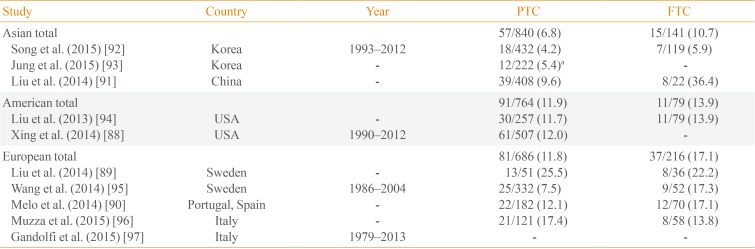
The prevalence of TERT promoter mutation exhibits significant variability among countries ranging from 4.2% to 25.5% [8992] of PTC and 5.9% to 36.4% [9192] of FTC (Table 5) [88899091929394959697]. Among these, the Korean prevalence was noticeably lower than other countries. We analyzed 551 patients with DTC in our institution, with TERT promoter mutations identified in 4.5% of patients [92]. Among 222 DTCs treated at the Catholic University of Korea, the overall prevalence of TERT promoter mutations was 5.4% [93]. The relatively large proportion of small-size tumors in Korea may account for the low frequency of these mutations relative to other countries. TERT promoter mutation assays are difficult to use in routine prognostic testing of DTC, especially in areas where its prevalence is low. Therefore, further studies identifying an optimal subset of TERT promoter mutations may be warranted.
CONCLUSIONS
Recent advances in molecular diagnostics have led to significant insights into the genetic basis of thyroid tumorigenesis, including a number of genetic alterations involved in the development of follicular cell-derived cancers having been reported. The frequency of each of these mutations varies significantly among populations, with Asian residents exhibiting significantly different mutational profiles relative to Western countries. Korean populations often exhibit different mutation rates relative to other countries, with BRAF mutation rates higher than any other country, whereas RET/PTC and PAX8/PPARγ rearrangements, and TERT promoter mutations, are generally lower. Awareness of the role and prevalence of each mutation may be important for the design of future studies, and may hold promise as either a diagnostic tool or a therapeutic target.
 XML Download
XML Download