

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
After the idea of intestinal factors regulating the function of endocrine secretion from the pancreas was first described by Moore and his colleagues in 1906, the incretins were introduced in the 1930s [1,2]. Incretins are intestinal hormones that regulate glucose-dependent insulin secretion in response to nutrient ingestion. Glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide are incretins [2]. GLP-1 is a 30-amino acid peptide secreted by intestinal L-cells [3] that regulates insulin secretion through binding to its receptor, GLP1R [4]. It is well established that GLP-1 augments insulin secretion in animal models and patients with type 2 diabetes [5,6]. GLP-1 stimulates insulin biosynthesis and growth, differentiation, and survival of β-cells [7,8,9,10]. In addition, GLP-1 decreases glucagon secretion on pancreatic β-cells [11]. In the brain, GLP-1 is known to promote satiety by regulating appetite and food intake, resulting in loss of body weight [12]. These diverse physiological functions of GLP-1 contribute to regulation of normal blood glucose level. The advantage of GLP-1 as a drug for type 2 diabetes is that it does not cause hypoglycemia under conditions of normal plasma glucose concentration due to its glucose-dependent action [11,13]. GLP-1 is able to stimulate insulin secretion in patients resistant to sulfonylurea drugs [14]. However, GLP-1 has an extremely short plasma half-life due to rapid degradation by dipeptidyl peptidase-IV (DPP-IV) [15]. DPP-IV selectively cleaves peptides with Pro and Ala in the second position. Exendin-4, which is found in the venom of the Gila monster, Heloderma suspectum, is a more potent GLP1R agonist than GLP-1 and is used clinically for treatment of type 2 diabetes [16,17]. Exendin-4 consists of 39 amino acids and exhibits 53% amino acid sequence similarity with GLP-1 [16]. The exendin-4 C-terminal tail, which consists of nine amino acid residues, enhances the binding affinity of exendin-4 to the N-terminal extracellular domain (ECD) of GLP1R [18]. Exendin-4 has a Gly residue in the second position, making it resistant to DPP-IV-mediated degradation [19]. However, exendin-4 is recognized as an antigen by the human immune system due to differences in amino acid sequence compared with GLP-1 [20]. Thus, more potent and long-acting GLP1R agonists are needed.
The amino acid sequence of GLP-1 is highly conserved among mammals and exhibits a high degree of identity among lower vertebrates [2,21,22,23]. It has been reported that non-mammalian GLP-1s are able to activate human GLP1R with high potency. Introduction of amino acid residues present in nonmammalian GLP-1 into human GLP-1 is a useful tool for developing novel GLP-1 analogs that are more potent than GLP-1. In the present study, we designed GLP-1 analog based on the amino acid sequences of Xenopus GLP-1 (xGLP) [24] and exendin-4. This analog exhibits resistance to DPP-IV mediated degradation and shows more potent activity toward GLP1R than wild-type human GLP-1.
METHODS
Peptides
GLP-1, exendin-4, xGLP, and the chimeric xGLP-E4 peptide were synthesized by AnyGen (Gwangju, Korea). The amino acid sequences of native GLP-1, exendin-4, xGLP, and xGLP-E4 are shown in Table 1.
Plasmid
The CRE-luc vector, which contains four copies of the cyclic AMP-responsive element (CRE: TGACGTCA) was from Stratagene (La Jolla, CA, USA). GLP1R cDNA was kindly provided by Dr. Bernard Thorens (Institute of Pharmacology and Toxicology, Lausanne, Switzerland).
Cell culture
HEK 293T cells were maintained at 37℃ in Dulbecco's Modified Eagle's medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS). INS-1 cells, rat pancreatic β-cells, were grown in RPMI 1640 medium containing 10% FBS at 37℃. Mouse pancreatic cells, β-TC-6 cells, were cultured in DMEM in the presence of 15% FBS at 37℃.
Transient transfection and luciferase assay
HEK 293T cells were plated in 48-well plates and transfected with Effectene reagent (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Approximately 48 hours after transfection, cells were treated with the respective ligands for 6 hours. Cells were then harvested, and luciferase activity was determined in cell extracts using a luciferase assay system according to standard methods for the Synergy 2 Multi-Mode Microplate Reader (BioTek, Winooski, VT, USA).
Stability of GLP-1 analogs
To evaluate the stability of GLP-1 and its analogs against DPP-IV activity, peptides were incubated at an initial concentration of 100 nM in DMEM containing 10% FBS or in 100% FBS at 37℃ for 0, 12, 24, 48, 72, and 96 hours in the presence or absence of 0.2 mM DPP-IV inhibitor diprotin A (Sigma-Aldrich, St Louis, MO, USA). FBS contains innate DPP-IV activity [25,26]. Residual peptide activity was then assessed by measuring luciferase activity after treatment of the GLP1R-and CRE-luc-cotransfected cells.
Insulin secretion
INS-1 cells and β-TC-6 cells were cultured in 24-well plates until 90% confluence. Cells were washed and incubated in KRB buffer (118.5 mM NaCl, 4.8 mM KCl, 2.7 mM CaCl2, 1.2 mM KH2PO4, 1.1 mM MgSO4·7H2O, 25 mM NaHCO3, and 4% bovine serum albumin, pH 7.4) and treated with low (2.8 mM) or high (16.7 mM) glucose for 2 hours at 37℃. Insulin secretion in response to treatment with 10 nM GLP-1 analogs was determined, after another 2-hour incubation of cells in KRB buffer containing low and high glucose, using an enzyme-linked immunosorbent assay kit according to the manufacturer's instructions (Millipore, Billerica, MA, USA).
Cell growth
INS-1 cells were seeded in 12-well plates at a density of 4×104 cells/well. At seeding and every 2 days thereafter, the respective peptides were added to the cells in fresh medium containing the appropriate concentration of glucose. Six and 10 days after cell seeding cells were washed with phosphate-buffered saline and detached with trypsin-ethylenediaminetetraacetic acid (Sigma-Aldrich). Harvested cells were counted with a hemocytometer.
RESULTS
Design of a chimeric GLP-1 analog
To develop a potent and long-acting GLP-1 analog, we performed amino acid sequence alignment of human GLP-1, xGLP, and exendin-4. These peptides share a high degree of amino acid sequence similarity at their N-terminal parts. Exendin-4 has a Gly residue in the second position and an additional nine residues at the C-terminal tail compared with human and xGLP (Table 1). This C-terminal tail is known to enhance the binding affinity of exendin-4 to the ECD of GLP1R [18]. We engrafted the nine C-terminal residues onto the C-terminal end of xGLP to increase the potency toward GLP1R. The Ala residue in the second position of xGLP was replaced with Ser to prevent DPP-IV mediated degradation.
Potency of xGLP-E4 toward GLP1R
The potency of xGLP-E4 was examined by luciferase assay in GLP1R and CRE-luc cotransfected HEK293T cells. The potency of exendin-4 (log EC50 value, -9.71±0.21) was slightly stronger than that of human GLP-1 (log EC50 value, -9.20±0.11) and xGLP (log EC50 value, -9.08±0.12). xGLP activated human GLP1R with potency equal to that of human GLP-1. xGLP-E4 exhibited higher activity (log EC50 value, -9.81±0.17) toward GLP1R than that of native human GLP-1 (Fig. 1).
Stability of GLP-1 analog
The stability of GLP-1 analogs was evaluated after incubation in DMEM containing FBS, which has innate DPP-IV activity. This assay was designed to measure bioactivity of the residual peptide to activate GLP1R in HEK 293T cells cotransfected with GLP1R and CRE-luc. When GLP-1 was incubated with 10% FBS, luciferase activity rapidly decreased. However, when diprotin A, a specific DPP-IV inhibitor, was added at the beginning of the incubation in the presence of FBS, the activity of GLP-1 was maintained for more than 72 hours (Fig. 2A). This result suggests that decreased activity of GLP-1 after incubation with FBS was due to degradation by DPP-IV. xGLP-E4 remained stable for more than 72 hours in DMEM containing 10% FBS, as did exendin-4 (Fig. 2A). xGLP-E4 and exendin-4 sustained more than 50% of their initial activity after 96-hour incubation with 100% FBS (Fig. 2B). GLP-1, however, completely lost its ability to activate GLP1R 72 hours after incubation in 100% FBS (Fig. 2B). These results show that xGLP-E4 is more stable in serum than wild-type human and xGLP.
Induction of glucose-dependent insulin secretion by xGLP-E4
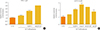
To investigate the effect of xGLP-E4 on insulin secretion, glucose-dependent insulin secretion was measured in rat (INS-1) and mouse (β-TC-6) pancreatic β-cells. In INS-1 cells, xGLP-E4 induced a higher level of insulin secretion than human GLP-1 in the presence of a high (16.7 mM) concentration of glucose (Fig. 3A). xGLP-E4 significantly increased insulin release at a high concentration of glucose in β-TC-6 cells (Fig. 3B).
Induction of β-cell proliferation by xGLP-E4
It is well known that GLP-1 increases β-cell proliferation [10]. To evaluate the effect of GLP-1 and analogs on β-cell growth, cells were treated with peptides every other day in the presence of low and high concentrations of glucose, and cells were counted 6 and 10 days after seeding. Under conditions of low glucose concentration, GLP-1 did not increase β-cell growth. However, exendin-4 and xGLP-E4 significantly stimulated β-cell proliferation 6 days after seeding (Fig. 4A). Ten days after cell seeding, GLP-1, exendin-4, and xGLP-E4 significantly increased β-cell proliferation. The effect of exendin-4 and xGLP-E4 on β-cell growth was greater than that of GLP-1 (Fig. 4B). It is well known that high concentration of glucose promotes cell proliferation and growth [27]. Under high glucose condition, the effect of glucose on cell growth is likely greater than that of GLP-1 analogs. Thus, growth effect of GLP-1 analogs may not be seen under high glucose concentration condition (Fig. 4). However, 10 days after cell seeding, only xGLP-E4 significantly increased β-cell growth (Fig. 4B).
DISCUSSION
As GLP-1 stimulates insulin secretion and inhibits glucagon secretion in type 2 diabetes patients with residual β-cell function, the use of GLP-1 has been proposed as a novel therapeutic option. The short biological half-life, however, considerably limits the use of GLP-1 in diabetes therapy [28]. Therefore, two strategies have been pursued, the development of inhibitors of DPP-IV and DPP-IV-resistant analogs of GLP-1 [29,30]. Administration of DPP-IV inhibitors reduces glycemic excursion, enhances insulin secretion, and inhibits degradation of GLP-1 in normal and diabetic rodents, as well as in human subjects [30,31]. Experiments using these DPP-IV inhibitors in animals showed that GLP-1 degradation could be prevented, and glucose tolerance was improved [31,32]. However, because DPP-IV ubiquitously degrades important regulatory peptides in addition to GLP-1 [33], the use of a DPP-IV inhibitor might theoretically cause widespread deleterious biological effects. The strategy of using GLP-1 analogs for diabetes therapy has the advantage over pharmacological DPP-IV inhibition that it is highly specific and does not lead to ubiquitous inhibition of the degradation of multiple regulatory peptides. Therefore, there has been much interest in designing more effective and long-acting GLP-1 analogs.
His1, Gly4, Thr7, and Asp9 in the N-terminal portion of GLP-1 are important for receptor binding and activation [34,35,36]. The hydrophobic face of GLP-1, formed by the conserved residues Phe22, Ile/Val23, Trp25, and Leu26, is responsible for interacting with the hydrophobic binding pocket in the ECD of GLP1R [37]. These critical amino acid residues are conserved between human and xGLP. For this reason, xGLP is able to activate human GLP1R with high potency (Fig. 1). The C-terminal region of exendin-4 binds to the N-terminal domain of the receptor and provides an additional interaction [18]. We generated xGLP-E4 having the C-terminal tail of exendin-4. xGLP-E4 showed higher activity toward GLP1R than wild-type human GLP-1. In addition, the stability and half-life of a drug are important prerequisites for clinical application. Human and xGLP have Ala at the second position, which is a target site of DPP-IV mediated degradation [38]. To prevent this degradation, we replaced Ala of xGLP-E4 with a Ser residue. The C-terminal tail derived from exendin-4 also protects this peptide from DPP-IV activity. As a result, the longevity of xGLP-E4 was extended, as was that of exendin-4.
The most important effect of a GLP-1 analog is the ability to stimulate glucose-dependent insulin secretion in β-cells. xGLP-E4 increased insulin secretion in rat and mouse β-cell lines at a high glucose concentration. Because xGLP-E4 did not increase insulin secretion at a low glucose concentration, long-term exposure of xGLP-E4 may not cause hypoglycemia. xGLP-E4 also induced β-cell growth, suggesting that this analog may be useful for treatment of type 1 diabetes. Currently, exendin-4 is clinically used for treatment of type 2 diabetes. Many studies have investigated the safety of it, and no evidence for cardiovascular, pulmonary, hepatic, or renal toxicity of exendin-4 has been reported [17,39]. The incidence of serious and severe adverse events is low although the most frequent adverse events are gastrointestinal symptoms, particularly nausea, vomiting, and diarrhea [17,39]. Since the amino acid sequence of xGLP-E4 is more similar to that of natural GLP-1 than exendin-4, xGLP-E4 may have fewer side effects when it is applied in vivo.
In summary, this study shows that modification of the amino acid sequence of native GLP-1 generates a novel GLP-1 analog with greater receptor affinity, biological activity, and stability. This analog may be a useful therapeutic agent for treatment of type 1 and 2 diabetes.




 XML Download
XML Download