

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Many studies have indicated a strong correlation between traumatic events during early life and the development of behavioral and neuroendocrine abnormalities later in life. Neonatal maternal separation is considered an animal model of stressful experience early in life. A number of studies have demonstrated its impact both on the development of depression [1,2] and anxiety-like behavior [3,4] later in life and on the activity of the hypothalamic-pituitary-adrenal (HPA) axis; i.e., permanent alterations in the characteristics of the HPA response to stress [5,6,7]. We have reported that repeated maternal separation during the first 2 weeks of life results in the development of depression- and/or anxiety-like behaviors in later life of rats with dysfunctions in HPA axis activity [8].
A recent human study reported that subjects offered palatable foods, such as savory snacks, bakery snacks and pasta, had more pleasant emotions such as satisfaction, enjoyment, and desire [9]. Exposure to a highly preferred diet high in fat was suggested to reduce stress sensitivity [10]. Earlier studies reported that rats consuming a high fat diet for a period of 4 months show decreased sympathetic responses following psychological and immunological stress [11], and that consumption of palatable food improves the stress response in rats subjected to restraint relative to rats fed chow only [12]. Another study showed that rats fed a high fat diet for 5 days are less anxious on the elevated plus maze compared to rats fed a low fat diet [13]. Studies have suggested that the HPA axis dysfunction is implicated in the pathophysiology of anxiety [14] and depression [15]. Taken together, these reports support the relationship between consumption of a highly palatable diet and positive emotional behavior, likely by a modulation effect of palatable food on the HPA axis function.
Here, we have hypothesized that psychoemotional disorders such as anxiety and depression induced by experience of neonatal maternal separation, which is accompanied by HPA axis dysfunction [16,17], is likely to be improved by consumption of highly palatable food (HPF) in later life. In this study, rat pups experienced maternal separation (MS) during the first 2 weeks after birth, and then had free access to Oreo cookies, a HPF rich in fat, in addition to standard chow during adolescence and youth. Anxiety- and depression-like behaviors and their HPA axis function responding to stress were examined at youth, and compared with nonhandled (NH) and MS control groups that were fed standard chow only.
METHODS
Animals
Sprague-Dawley rats were purchased (Samtako Bio, Osan, Korea), and cared in a specific-pathogen-free barrier area with constant control of temperature (22℃±1℃), humidity (55%), and a 12/12 hours light/dark cycle (lights-on at 7:00 AM). Standard laboratory food (Purina Rodent Chow, Purina Co., Seoul, Korea) and membrane filtered purified water were available ad libitum. Animals were cared according to the Guideline for Animal Experiments, 2000, edited by the Korean Academy of Medical Sciences, which is consistent with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals, revised in 1996. All animal experiments were approved by the Committee for the Care and Use of Laboratory Animals at Seoul National University.
Experimental protocol
Nulliparous females and proven breeder males were used for breeding in the laboratory of the animal facility, and the pups were reared in a controlled manner to minimize and standardize unwanted environmental stimulation from in utero life. Twelve hours after confirming delivery (postnatal day [PND] 1), pups were culled to five males and five females per litter. Each litter was assigned either for the MS group or for the NH group. MS was performed as we previously described [16,17,18,19]. In brief, MS pups were removed from their dam and home cage and placed closely together in a new cage bedded with woodchips (Aspen shaving, Animal JS Bedding, Cheongyang, Korea) for 180 minutes, and then returned to their home cage and dam. MS was performed at room temperature; i.e., no additional treatment to keep the pups warm during the separation period other than placing them closely together was offered and pup-cooling during MS was expected. MS was performed from 9:00 AM to 12:00 PM daily from PND 1 through 14, and then the pups were left with their dam undisturbed until weaning on PND 22. The NH group remained undisturbed until weaning except for routine cage cleaning. For cage cleaning, all rats were moved to clean the cage twice a week. On the weaning day, two NH and four MS male pups were randomly selected from each NH or MS litter and placed two NH or two MS pups together in each cage. Two MS pups housed together from each litter received free access to chocolate cookies, a HPF, in addition to ad libitum chow from PND 28 (maternal separation fed with chow and cookies [MS+HPF] group), and the remaining two MS pups in each litter (MS group) and NH pups (NH group) received standard chow only. Water was freely available for all of the experimental groups, and the food conditions were continued throughout the whole experimental period.
Ambulatory activity
NH, MS, and MS+HPF rats were subjected to the ambulatory test on PND 54. On each trial, the rat was placed in the center of the activity chamber (43.2 cm in length, 42.2 cm in width, and 30.5 cm in height, MED Associates, St. Albans, VT, USA), a transparent acryl chamber equipped with two horizontal planes of 16 infrared photocell-detector pairs placed in the x, y dimension, spaced 2.5 cm apart, and its ambulatory activity was monitored by the computerized system for 30 minutes. The light condition of the test room was maintained at the same intensity as the animal rooms under day light condition. Ambulatory activity was measured as the total counts of beam interruptions in the horizontal sensor during each consecutive 5 minutes session. Defecation activity of each rat during the ambulation test was measured by the weight of the fecal boli. Grooming activity was further analyzed; i.e., forepaw and head grooming was considered as rostral grooming, and body, legs, and tail/genital grooming as caudal grooming [20]. The activity chamber was cleaned with 70% ethanol after each use to eliminate any olfactory cues of the previously tested rat.
Elevated plus maze
Two days after the ambulatory activity test (PND 56), rats were subjected to the behavioral assessment in an elevated plus maze, a plus shaped acryl maze with two opposite open arms (50 cm in length and 10 cm in width) and two opposite closed arms (50 cm in length, 10 cm in width, and 31 cm in height), extending out from a central platform (10×10 cm). The whole apparatus was elevated 50 cm above the floor. The test procedure was followed as previously described [4]. Each rat was placed in the center of the maze facing one of the open arms, and then allowed to explore the open or closed arms of the maze for 5 minutes. The time spent in the different arms was recorded, respectively. Four paws had to be inside the entrance line to each arm, which signaled the start of the time spent in the specific arm, and the end time was recorded when all four paws were outside the line again. The maze was cleaned with 70% ethanol after each test to prevent influences of the previously tested rat.
Forced swim test
Three days after the elevated plus maze test (PND 59), rats were subjected to the forced swim test, according to the method previously described [21]. Each rat was allowed to swim in a glass cylinder (54 cm in height and 24 cm in diameter) filled with water in 40 cm of depth (23℃ to 25℃) for 5 minutes, and the test sessions were recorded by a video camera from the side of the cylinder. Duration of rat's immobility in the water was scored from videotapes by a trained observer (J.Y.K.) who was blinded to the experimental conditions. Immobility was defined as the state in which rats were judged to be making only the movements necessary to keep their head above the surface.
Plasma corticosterone assay
A week after the end of behavioral sessions, rats were placed in a restraint box for 2 hours, in which rats were able to move their four limbs but not to change their body orientation. Tail blood was collected at 0, 30, 60, and 120 minutes time points during the restraint period, and centrifuged at 2,000 rpm for 20 minutes. The plasma samples were frozen in liquid nitrogen, and stored at -80℃ until used for the assay. Plasma levels of corticosterone were determined by radioimmunoassay using 125I-labelled Coat-A-Count kit (Siemens, Mountain View, CA, USA). The sensitivity of the assay was 5.7 ng/mL. The intra-assay coefficient of variation was 4% to 12.2%.
Rats were placed in the test room at least 2 hours prior to each test to minimize unwanted stress effects, and all behavioral assessments were performed between 9:00 AM and 12:00 PM of the day to avoid the influences of circadian variances.
Statistical analysis
Data were analyzed by one-way analysis of variance, and preplanned comparisons between groups performed by post hoc Fisher's protected least significant difference test, using StatView software (Abacus, Berkeley, CA, USA). The level of significance was set at P<0.05, and all values were presented as means±SEM.
RESULTS
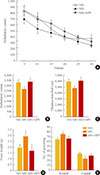
Daily chow intake and body weight gain did not differ between NH and MS rats (Fig. 1A, B). Chow intake of MS rats was decreased with cookie access, but daily caloric intake was not affected (Fig. 1A). Consequently, body weight gain of MS rats was not affected by cookie access (Fig. 1B).
Ambulatory activities of NH, MS, and MS+HPF rats were measured in a computerized activity chamber for 30 minutes on PND 54. Ambulatory counts, the total counts of beam interruptions in the horizontal sensor, were gradually decreased during the test session in all groups (Fig. 2A). Total ambulatory counts and the distance travelled during the 30 minutes of session were decreased in MS rats compared with NH rats (P<0.05), and increased with cookie access in MS rats (MS vs. MS+HPF, P<0.05) (Fig. 2B, C). Defecation activity and grooming behavior was scored during the ambulatory activity test (Fig. 2D, E). Feces weight and rostral grooming were increased whereas caudal grooming decreased by MS experience (NH vs. MS, P<0.05), and these behaviors were normalized with cookie access.
In order to further assess the anxiety-like behaviors, rats were subjected to an elevated plus maze test 2 days after the ambulatory activity test. MS rats spent more time in closed arms, and less time in open arms, compared with NH rats (P<0.05), and cookie access did not affect the behavioral scores of MS rats (Fig. 3A). To assess depression-like behaviors, rats were subjected to forced swim test 3 days after the elevated plus maze test. Immobility duration during the 5 minutes of test session was significantly increased (P<0.05) in MS rats (27.667±5.878 seconds) compared with NH rats (8.222±2.748 seconds), and the immobility score was not affected by cookie access in MS rats (Fig. 3B).
A week after the swim test, rats received 2 hours of restraint stress, and tail blood was collected at 0, 30, 60, and 120 minutes time points during the restraint session, and used for plasma corticosterone assay (Fig. 4). The plasma corticosterone levels of NH rats were significantly increased at 30 and 60 minutes after the onset of the restraint stress (P<0.05 vs. 0 time point), and restored to the basal levels at 120 minutes after the restraint onset. However, in MS rats, stress-induced corticosterone increase was not observed, and the basal levels of corticosterone (0 time point) were higher than in NH rats (138.695±28.892 ng/mL in NH and 236.143±36.524 ng/mL in MS, P<0.05). MS rats with cookie access (MS+HPF) showed a significant increase in plasma corticosterone levels at 30 minutes time point (P<0.05 vs. 0 time point), and their basal plasma corticosterone levels did not differ from NH rats.
DISCUSSION
Many studies have demonstrated that experience of neonatal MS may lead to the development of depression- and anxiety-like behaviors later in life [2,3,4,5]. In this study, not only the ambulatory activities but also the distance travelled during the activity test were suppressed significantly in MS rats fed with chow only compared with NH rats, suggesting increased anxiety and/or depression by experience of our MS protocol. Defecation scores during the activity test were increased in MS rats fed with chow only relative to NH rats in this study. Low locomotor activity and high defecation rates are often regarded as central emotional responses of rats exposed to novelty [22]. Grooming is often seen in animal models of stress and anxiety [23,24], leading to a long-standing view of grooming as an anxiogenic response [25,26]. It has been reported that highly stressed mice spend significantly more time grooming rostral than caudal areas [20,27]. In this study, MS rats fed with chow only spent more time in rostral grooming and less time in caudal grooming, compared to NH rats. That is, increased defecation scores and rostral grooming, and decreased caudal grooming in MS rats fed with chow only further supports the anxiogenic effect of MS in our protocol. It has been shown that alterations in the depression-like states in rodents by chronic stress affect grooming behaviors [28,29,30]. Thus, it is likely that altered grooming behaviors in our MS rats fed with chow only may also reveal increased depression-like states. Indeed, MS rats fed with chow only showed behavioral depression in the swim test with increased immobility duration, and increased anxiety in the elevated plus maze test, compared to NH rats. Together, it is concluded that the rats subjected to our MS protocol show anxiety- and depression-like behaviors in later life, in accordance with our previous reports [17,19].
It has been reported that MS experiences during neonatal period results in permanent alterations in the characteristics of the HPA response to stress [5,6,7]. In this study, MS rats fed with chow only showed blunted corticosterone response to restraint stress with increased basal levels of corticosterone compared with NH control rats. This result concurs with our previous report showing altered HPA axis activities in our MS rats responding to metabolic stresses [16,18], and further supports the HPA axis dysfunction by experience of our MS protocol. It is likely that increased basal corticosterone levels in MS rats fed with chow only may reveal increased HPA axis activity responding to daily stressors by MS experience and result in a blunted response to acute restraint stress. Studies have suggested that the HPA axis dysfunction is implicated in the pathophysiology of anxiety [14] and depression [15]. Elevated basal cortisol levels were associated with anxiety and depressive symptoms in young subjects [31,32,33]. Thus, it is plausible that the HPA axis dysfunction in our young MS rats; i.e., blunted stress-induced corticosterone increase with elevated basal levels, is implicated in their anxiety- and depression-like behaviors observed.
In this study, MS rats, that had free access to chocolate cookies in addition to the chow access (MS+HPF group), showed a significant increase in the plasma corticosterone levels responding to restraint stress, unlike the blunted response in MS rats fed with chow only (MS group). Furthermore, the basal corticosterone levels of MS+HPF rats did not differ from NH rats. These results suggest that free access to HPF during adolescence and youth might have improved the HPA axis dysfunction by MS experience. Puberty is a crucial developmental period characterized by increased endocrine plasticity and changes in stress responsiveness [34]. Studies comparing HPA axis activity in prepubertal and adult rats show that there is a significantly persistent hormonal stress response in prepubertal animals compared to adults [35,36,37]. Previous study suggested that a postweaning highly preferred diet can modify the basal HPA axis activity and the endocrine responses to an acute stress by affecting both stress and metabolic mediators [38,39]. Exposure to a highly preferred diet high in fat was suggested to reduce stress sensitivity [10]. Earlier studies reported that sympathetic responses to psychological stress are decreased in rats consuming a high fat diet for a long-time period [11], and that consumption of palatable food improves the stress response in rats subjected to restraint [12]. Also, exaggerated corticosterone release following acute stress was observed in a study using a 7-day moderate high fat diet (20% fat content) protocol to dissociate the impact of obesity from that of fat [40]. The chocolate cookies used in this study is a moderate high fat diet (~21% fat content), and caloric intake and body weight gain of MS rats was not affected by cookie access. Altogether, it is concluded that free access to a highly preferred diet during adolescence and youth may reduce the stress sensitivity of MS rats to daily routine stressors and normalize the basal corticosterone levels which were increased by MS experience. Consequently, MS rats fed with HPF were likely able to show a normal stress response; i.e., a significant increase of corticosterone levels in response to an acute restraint stress.
In this study, ambulatory activities and the distance travelled during the activity test were significantly increased, and defecation activity decreased, in the MS+HPF group compared to the MS group. Also, grooming behaviors of MS+HPF rats did not differ from NH rats, while rostral grooming was increased and caudal grooming decreased in MS rats compared with NH rats. Previous studies showed that prolonged fat intake decreased anxiety-like behaviors with reduced sympathetic response to stressors [11] and short-term exposure to a high fat diet also reduced anxiety on the elevated plus maze in rats [13]. Together, it is suggested that prolonged access to chocolate cookies, a moderate high fat diet, during adolescence and youth might have improved anxiety-related behaviors of MS rats in this study. As mentioned above, HPA axis dysfunction has been implicated in the pathophysiology of anxiety and depression [14,15], and effective treatments of affective disorders with antidepressants or anxiolytic agents also modulate the HPA axis function [41,42]. In this study, although some anxiety-related behaviors of MS rats and their HPA axis functions were improved together by prolonged cookie access, anxiety on the elevated plus maze and depressive behavior during the swim test were not improved. Thus, it is concluded that prolonged cookie access during adolescence and youth partly improves anxiety-related, but not depressive, symptoms in rats that experienced our MS protocol, possibly in relation with improved HPA axis function.
The nucleus accumbens (NAc), a basal forebrain structure constituting a mesolimbic dopaminergic pathway, has a role in reward, motivation, and reinforcement [43]. It has been reported that reduced dopaminergic function within the NAc may cause depression-like behaviors in rodents [44], and the striatal dopaminergic activity was suggested to be associated with the severity of anhedonia in depressed patients [45]. The NAc is activated in response to behavioral stress paradigm [46,47], suggesting its implication in the stress-responsive HPA axis regulation. Rats that experienced our MS protocol showed not only HPA axis dysfunction but also reduced dopaminergic activity in the NAc [48]. Consumption of palatable foods elevates extracellular dopamine concentrations in the NAc of the ventral striatum [49], and the striatal dopamine release reflects the perceived pleasantness of a meal in humans [50]. However, we have found that limited cookie access during adolescent period did not affect the NAc dopamine levels in MS rats, but normalized the amygdala dopamine levels which was increased by MS experience (our unpublished observation). Mesolimbic dopamine neurons originating in the ventral tegmental area (VTA) innervate limbic sites including the NAc and amygdala. The VTA, Nac, and amygdala are posited to form an interconnected network important for reward processing [51], while the amygdala also has a fundamental role in the control of anxiety [52,53]. The NAc has been viewed as a transition area between the ventral striatum and the extended amygdala including the central nucleus of the amygdala (CeA) [54]. Studies have suggested that enhanced dopamine signaling within the CeA is associated with reward-seeking [55,56] and anxiety-like behaviors [57,58]. The CeA has been shown to respond to hormonal changes associated with stress and fear, the core symptoms of anxiety disorders [59], and interact with the HPA axis [60,61]. Taken all together, it is likely that altered dopaminergic activity in the NAc and CeA with the HPA axis dysfunction may be implicated in MS-induced depression- and anxiety-like behaviors, and speculated that the partial improvement of anxiety-related behaviors in MS rats by prolonged palatable food consumption may be due to a region specific effect of it on brain dopamine neurotransmission. Further studies are warranted to define its underlying mechanism.



 XML Download
XML Download