

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In inflammatory conditions, a syndrome referred to as the sickness response occurs in a number of different ways. Reduced appetite and body weight loss are typical symptoms of inflammatory diseases [1-3]. Anorexia, a symptom of decreased appetite, is associated with a wide range of acute and chronic pathological conditions which are characterized by an inflammatory state, such as bacterial or parasitic infection, cancer or chronic inflammatory bowel disease [4,5]. This sickness-related anorexia negatively affects recovery and treatment success with a decreased quality of life. A prolonged reduction in food intake will deplete body fat and protein reserves, resulting in malnutrition, impaired host immune functions, and increased morbidity and mortality [6]. Therefore, elucidating the mechanism of infection-associated anorexia and weight loss is essential to improve patient quality of life.
Proinflammatory cytokines play critical roles in the etiology of disease- and infection-related anorexia [7-9]. The brain is an important target for proinflammatory cytokines that affect the neuronal circuits controlling food intake and energy homeostasis [10,11]. Inflammation in the brain results in a profound state of negative energy balance that is an adaptive response to infection [12,13]. Administration of bacterial endotoxins, such as lipopolysaccharide (LPS), has been demonstrated to induce sickness response behavior in laboratory animals by inducing acute inflammation in the brain as well as the peripheral organs [14,15].
Members of the tumor necrosis factor receptor superfamily (TNFRSF) play a key role in regulating the immune response to infection [16]. Herpes virus entry mediator (HVEM, also called as TNFRSF14) is a membrane receptor that binds several ligands and mediates costimulatory pathways, predominantly through its interaction with LIGHT (also called as TNFSF14). HVEM also mediates coinhibitory effects, predominantly by interacting with immunoglobulin (Ig) superfamily members, B- and T-lymphocyte attenuator, or CD160 [17]. Therefore, HVEM can be considered a molecular switch for various activating or inhibitory functions [18,19].
HVEM activates the nuclear factor (NF)-κB pathway, leading to the induction of proinflammatory genes [20]. Recently, several studies have reported that HVEM-mediated stimulatory signals are essential for promoting proinflammatory cytokine production and intestinal pathology [21,22] and are important in hepatic inflammatory disease [23]. Moreover, HVEM deficiency attenuates the inflammatory responses of adipose tissue and glucose intolerance in diet-induced obesity [24]. Collectively, these data implicate that HVEM plays a proinflammatory role in peripheral tissues. However, little is known about its function in the brain.
In this study, we assessed changes in food intake and body weight after intraperitoneal (IP) administration of LPS or intracerebroventricular (ICV) injection of LIGHT and demonstrated that HVEM deficiency ameliorated inflammation-induced anorexia and weight loss. These results indicate that HVEM may be a novel therapeutic target for inflammation-induced negative energy balance.
METHODS
Animal care and treatment
Eight- to 10-week-old male C57BL/6 mice and mice lacking HVEM expression (HVEM-/-) were maintained under specific pathogen-free conditions [25]. Mice were fed standard chow (Feedlab, Guri, Korea) ad libitum. Animals were housed under controlled temperature (23℃ to 25℃) and a 12-hour light-dark cycle, with light from 6:00 AM to 6:00 PM. Animal experiments were conducted in accordance with the regulations of University of Ulsan for the Care and Use of Laboratory Animals (Permission No. UOU-2011-005).
Mice were IP-injected with LPS (100 µg/kg, Sigma-Aldrich, St. Louis, MO, USA) or ICV-injected with recombinant mouse LIGHT (300 ng/2 µL, Research & Diagnostics, Minneapolis, NE, USA) at 4:00 PM. Food intake and body weight were measured at 24 hours after injection of LIGHT and LPS.
Stereotaxic surgery and ICV-administration
Mice were anesthetized with an IP-injection of tribromoethanol (250 µg/mouse, Sigma-Aldrich) and were placed in a stereotaxic apparatus (Stoelting, Wood Dale, IL, USA). A polyethylene cannula was implanted into the lateral ventricle (coordinates of 0.03 mm posterior, 0.1 mm lateral, and 2.4 mm ventral to the bregma) and secured to the skull with dental cement. Mice were immediately placed in individual cages. After 1 week of recovery, test materials were injected through the cannula. Recombinant mouse LIGHT (300 ng) was injected in 2 µL total volume.
Immunohistochemistry
The mice were anesthetized with tribromoethanol and transcardially perfused with 10 mL ice-cold 0.1 M phosphate buffer (PB, pH 7.4), followed by 10 mL 4% paraformaldehyde. The brains were dissected and postfixed overnight in the same fixative containing 20% sucrose. Brains were dehydrated and placed in an embedding medium (Tissue-Tek, Torrance, CA, USA), and slide-mounted sections were prepared with a cryostat. The frozen sections of brain were completely dried and washed with 0.1 M PB for 30 minutes. Sections were incubated with blocking buffer (3% nonfat milk, 0.3% Triton X-100 and 0.1 M PB). After washing the slides three times with 0.1 M PB for 5 minutes, slides were incubated with goat antimouse HVEM antibody (1:250, Santa Cruz, Billerica, MA, USA) overnight at room temperature and then washed three times with 0.1 M PB for 10 minutes. After washing, the sections were incubated with biotin-conjugated donkey antigoat IgG (1:400, Vector Labs, Burlingame, CA, USA) in PB for 2 hours. Staining was performed using an ABC kit (Vector Labs), color development was achieved by incubating the sections for 5 minutes with the tyramide signal amplification system (NEN Life Science, Boston, MA, USA), and images were obtained using fluorescence microscopy.
Statistical analyses
All results are expressed as mean±standard error of measure (n are given in the figure legends). Statistical analyses were performed using repeated measures analysis of variance with Bonferroni post hoc analyses. A two-tailed unpaired Student t test was used for comparison between two groups.
RESULTS
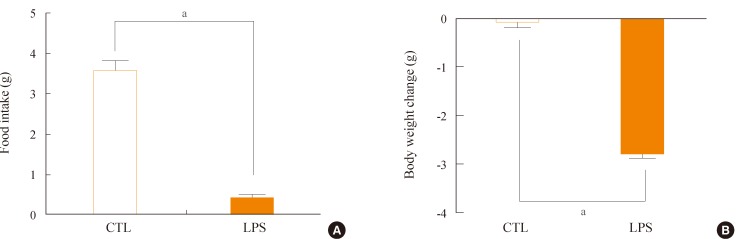
In order to assess role of LPS in energy homeostasis, mice were injected with LPS into the abdominal cavity. Administration of LPS significantly decreased food intake, measured for 24 hours after the injection of LPS (Fig. 1A). This anorectic effect of LPS resulted in a decrease of body weight over the 24 hours duration (Fig. 1B). Control saline-injected animals showed an average of 0.1 g of weight loss over the 24 hours, while LPS-injected mice lost an average of 2.78 g. These results suggest that the administration of LPS decreased food intake, thereby reducing body weight.
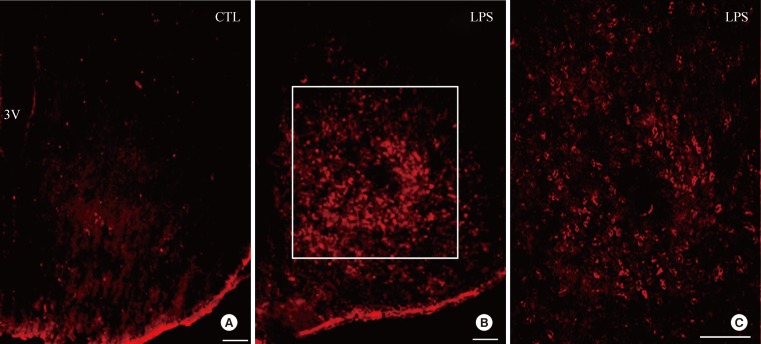
We next assessed if LPS induced change in the hypothalamic expression of HVEM using immunohistochemistry. IP-injection of LPS strongly increased the HVEM immunoreactivity in the hypothalamic arcuate nucleus (ARC) (Fig. 2), suggesting that the LPS-induced inflammation affects HVEM expression in the ARC.
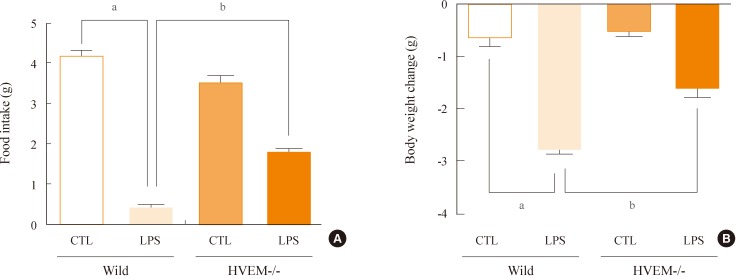
Because LPS increased the expression of HVEM in the ARC, we hypothesized that HVEM signaling might be required for the LPS-induced anorexia and body weight loss. To investigate the role of HVEM during the LPS-induced anorexia and body weight loss, the HVEM-/- mice were injected with LPS and food intake and body weight were measured. As already shown in Fig. 1, the wild-type mice exhibited a decreased food intake and body weight following the IP administration of LPS (Fig. 3). However, LPS induced only a minor decrease in food intake and body weight in the HVEM-/- mice (Fig. 3). These results suggest that the HVEM signaling plays a pivotal role in the LPS-induced anorexia and body weight loss.
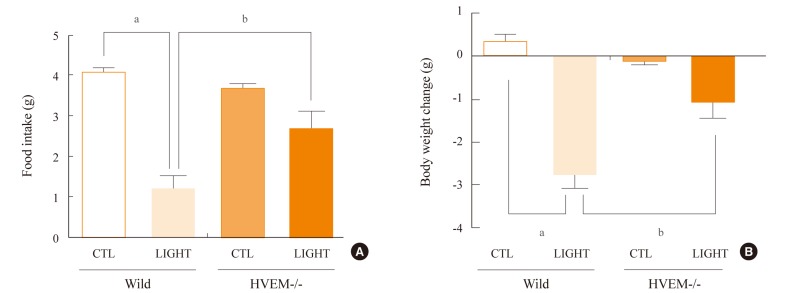
The interaction of HVEM with its ligand LIGHT has been known to promote inflammatory responses and the subsequent production of proinflammatory cytokines [26]. Thus, we examined the effect of LIGHT on the anorexia and body weight loss. Mice were ICV-injected into the lateral ventricle with recombinant LIGHT. The ICV administration of LIGHT significantly decreased food intake and body weight in the wild-type mice for 24 hours (Fig. 4). However, the same administration caused a significantly smaller decrease in food intake and body weight in the HVEM-/- mice during the same observation period (Fig. 4). These results suggest that the interaction of LIGHT with HVEM in the brain plays an essential role in the regulation of food intake and body weight.
DISCUSSION
Reduced appetite is a hallmark of the inflammatory diseases [1-3,27] and is concomitant with many chronic pathological conditions, including cancer. This negative energy balance is controlled by the proinflammatory cytokines in the brain [10,11]. Members of the TNFRSF play a key role in regulating immune responses to infection [16]. HVEM, a member of TNFRSF, activates transcription factors, such as NF-κB and AP-1 [20], which control the expression of genes involved in multiple immune, inflammatory, and acute-phase responses to infection [28]. Recent studies have demonstrated that HVEM signaling plays a critical role in promoting proinflammatory cytokine production in peripheral tissues, where cytokines are involved in intestinal inflammation, hepatitis, and diet-induced obesity [21-23].
In this study, we demonstrated that the HVEM signaling in the brain contributes to acute inflammation-induced anorexia and weight loss. At the beginning of this study, we found that severe anorexia and body weight loss was induced in mice suffering from intestinal inflammation caused by the administration of dextran sulfate sodium (data not shown). However, we found that the HVEM-/- mice showed attenuated intestinal inflammation and body weight loss (data not shown), as previously reported [21]. Therefore, the attenuated inflammation in HVEM-/- mice might contribute to the amelioration of anorexia. This study together with the previous report [21] indicates that the HVEM signaling is necessary for the intestinal inflammation-induced anorexia and body weight loss.
LPS is a representative bacterial endotoxin for the simulation of acute bacterial infection and inflammation. In this study, we found that administration of LPS significantly reduced food intake as well as body weight, and moreover, increased the expression of HVEM in the hypothalamic ARC. It has been reported that IP-injection of LPS strongly increased serum levels of proinflammatory cytokines, which affect the hypothalamic center for energy homeostasis [29]. The hypothalamus plays a crucial role in the control of feeding behavior and energy expenditure [30,31]. Numerous inflammatory stimuli activate the hypothalamic proinflammatory cytokines, such as tumor necrosis factor-α, interleukin-1β (IL-1β), and IL-6, which are involved in the sickness behavior characterized by anorexia and fever [32].
It is likely that the LPS-induced activation of hypothalamic HVEM signaling plays an important role in activating proinflammatory cytokines, subsequently resulting in the decreases of food intake and body weight. Indeed, loss of HVEM expression in the HVEM-/- mice (and thus, loss of the activation of its signaling) gave rise to the improvement from the LPS-induced anorexia and body weight loss. Peripheral inflammatory mediators stimulated by LPS may exert brain responses via HVEM signaling. Indeed, peripheral administration of LPS was shown to activate production of inflammatory cytokines that cause neuroinflammatory responses in the brain, particularly in the microglia and astrocytes [33]. HVEM might be induced in the microglia and astrocytes during this neuroinflammatory process as shown in this study.
To address whether the activation of brain HVEM is responsible for inflammation-induced negative energy balance, changes in food intake and body weight were measured in mice received LIGHT, a stimulatory ligand of HVEM, in their brain. ICV-administration of LIGHT significantly decreased food intake in the wild-type mice, suggesting that the activation of brain HVEM is a plausible cause of the inflammation-induced negative energy balance. Moreover, the ICV-injection of LIGHT was less effective in decreasing food intake and body weight in the HVEM-/- mice, further indicating that the activation of brain HVEM by LIGHT (or other inflammatory stimuli) is responsible for the negative energy balance. However, LIGHT-induced decreases in food intake and body weight were not completely suppressed in the HVEM-/- mice, likely owing to the fact that LIGHT can exert its effects through other receptors, such as the lymphotoxin β receptor [34].
Taken together, this study suggests that HVEM signaling in the brain plays an important role in regulating energy homeostasis by activating inflammation-induced anorexia and body weight loss and that HVEM is a good candidate for a therapeutic target to improve quality of life for inflammatory disease patients.
 XML Download
XML Download