

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
There are diverse types of ginseng, and it has been reported that some types improve blood glucose levels in patients with type 2 diabetes mellitus [1234]. Among them, Panax ginseng (Asian ginseng) and ginsam, a vinegar extract from P. ginseng, have been frequently reported to decrease blood glucose levels and body weights in animal models [567] and in humans [4]. Ginsenosides are biologically active components of P. ginseng, and there are many different types of ginsenoside with different pharmacological activities [8]. The ginsenoside Rg3, one of the most enriched components of ginsam, has been suggested as the major component producing anti-diabetic effects [6].
The main proposed mechanism of the anti-diabetic effect of P. ginseng is the improvement in insulin resistance in skeletal muscle rather than an increase in insulin secretion from pancreatic β-cells [56]. It has been demonstrated that Rg3 activates the phosphorylation of insulin receptor substrate-1 (IRS-1) and Akt, and glucose uptake in L6 myotubes [9].
Mitochondria play a pivotal role in the insulin resistance of skeletal muscle [1011]. Therefore, it is hypothesized that the improvement in insulin resistance by P. ginseng or Rg3 is, at least in part, mediated through the regulation of mitochondrial biogenesis or function in skeletal muscle. Here, we investigated the effects of Rg3 on the mitochondria of skeletal muscle using C2C12 myotubes.
METHODS
Cell culture
C2C12 myoblasts (ATCC, Manassas, VA, USA) were maintained in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA). Two days after the cells reached confluence, differentiation was induced with 1×high-glucose DMEM with 2% horse serum (Invitrogen) for 4 days. To induce insulin resistance, C2C12 myotubes were treated with 500 µM palmitate (Sigma-Aldrich Co., St. Louis, MO, USA) for 24 hours. To induce mitochondrial dysfunction, C2C12 myotubes were treated with 10 µM antimycin A (Sigma-Aldrich Co.) for 48 hours. After pretreatment with antimycin A, palmitate, or dimethyl sulfoxide (DMSO), 100 µM Rg3 (Sigma-Aldrich Co.) or DMSO was added to the medium for 24 hours. In some experiments, 10 µM compound C (Sigma-Aldrich Co.) was added 1 hour before Rg3 treatment.
Quantitative reverse transcription polymerase chain reaction
Total RNA from C2C12 cells was isolated using TRIzol (Invitrogen). Reverse transcription polymerase chain reaction (RT-PCR) was performed using SYBR-master mix (TaKaRa, Kyoto, Japan) and the ABI 7500 RT-PCR system (Applied Biosystems, Foster City, CA, USA). The primer sequences for RT-PCR are shown in Table 1. All data were analyzed using ABI 7500 software version 2.0.5. (Applied Biosystems).
Cellular adenosine triphosphate levels
C2C12 cells were homogenized in 200 µL of mammalian cell lysis solution (ATPlite; PerkinElmer Inc., Waltham, MA, USA) using a micro-homogenizer. The cell lysates were aliquoted into a 96-well microplate (50 µL per well); 50 µL of substrate solution (ATPlite) was added to each well. After a dark adaptation for 10 minutes, luminescence was detected on a microplate reader (VICTOR3; PerkinElmer Inc.).
Oxygen consumption rate
C2C12 cells were cultured in DMEM medium (Sigma-Aldrich Co.). The oxygen consumption rate (OCR) was measured using the Seahorse XF24 analyzer (Seahorse Bioscience Inc., North Billerica, MA, USA) according to the manufacturer's protocol. The OCR was measured under basal conditions and after the addition of the mitochondrial inhibitor oligomycin (2 µM) and the mitochondrial uncoupler carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP; 5 µM) to assess maximal oxidative capacity. Antimycin A and rotenone were then added to inhibit the mitochondrial complex completely.
Immunoblotting
C2C12 cells were lysed in 20 mM Tris-HCl (pH 7.4), 5 mM Na4P2O7, 100 mM NaF, 2 mM Na3VO4, and 1% NP-40 buffer supplemented with a protease inhibitor (1 µg/µL aprotinin, 1 µg/µL leupeptin, and 1 mM PMSF). Cell lysates were sonicated on ice twice for 15 seconds each time, and debris was removed by centrifugation (15,000 g) for 30 minutes at 4℃. Proteins (10 to 20 µg) were separated on a sodium dodecyl sulfate-polyacrylamide gel and transferred to a nitrocellulose membrane (Whatman, Dassel, Germany). The membrane was blocked with 5% skim milk in Tween-20-Tris-buffered saline (TBS-T) for 1 hour at room temperature, followed by incubation with primary antibody overnight at 4℃. The primary antibodies used were a total OXPHOS antibody cocktail (MitoScience, San Francisco, CA, USA), anti-IRS-1 (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), anti-pIRS-1 (Cell Signaling, Danvers, MA, USA), anti-Akt (Cell Signaling), anti-pAkt (Cell Signaling), anti-adenosine monophosphate (AMP)-activated protein kinase (AMPK; Cell Signaling), anti-phosphorylated AMPK (Cell Signaling), and anti-γ-tubulin (Sigma-Aldrich Co.). The secondary antibodies were anti-rabbit or anti-mouse immunoglobulin G.
RESULTS
Rg3 enhances the insulin signaling pathway
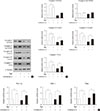
Insulin stimulated IRS-1 and Akt phosphorylation in C2C12 myotubes, and Rg3 treatment for 24 hours enhanced the insulin signaling pathway (Fig. 1). To induce insulin resistance, we pretreated the cells with palmitate, and it decreased IRS-1 and Akt phosphorylation. Under that condition, Rg3 treatment increased IRS-1 and Akt phosphorylation (Fig. 1A) Even after pretreatment with antimycin A, the oxidative phosphorylation complex III inhibitor, Rg3 increased IRS-1 and Akt phosphorylation (Fig. 1B), suggesting that Rg3 can improve insulin signaling which has been deteriorated by mitochondrial dysfunction.
Rg3 improves mitochondrial function
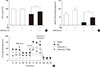
Next, we assessed the effects of Rg3 on mitochondrial function by measuring cellular adenosine triphosphate (ATP) levels or the OCR. Under basal conditions, there was no difference in cellular ATP levels after Rg3 treatment (Fig. 2A). However, after the induction of mitochondrial dysfunction by antimycin A, Rg3 significantly increased ATP production from 68% to 84% (P=0.03) (Fig. 2A). The effects of Rg3 on mitochondrial function were more pronounced when analyzing the OCR. The overall OCR after Rg3 treatment was increased (Fig. 2B). The OCR increase was clearly demonstrated under the conditions of antimycin A-induced mitochondrial dysfunction; the basal OCR was significantly increased by 2.8-fold (P=0.03) (Fig. 2C).
Rg3 increases the expression of key genes involved in mitochondrial biogenesis
Next, we evaluated the quantitative changes in the protein or mRNA levels of mitochondrial complexes. Both Western blotting and quantitative RT-PCR analysis revealed that Rg3 increased CoxII of complex IV and F1α of complex V, while the effects of Rg3 were minimal or inconsistent in the other mitochondrial complexes (Fig. 3A and B). Under antimycin A-induced mitochondrial dysfunction, similar effects were observed. Therefore, we next determined the expression of transcription factors which regulate the expression of mitochondrial proteins, using quantitative RT-PCR. The expression of peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α), nuclear respiratory factor 1 (NRF-1), and mitochondrial transcription factor (Tfam) was significantly increased after Rg3 treatment, regardless of the presence of antimycin A (P<0.05) (Fig. 3C). These results suggest that Rg3 enhances expression of mitochondrial proteins through an increase of these transcription factors, which leads to the promotion of mitochondrial biogenesis.
Rg3 is not mediated by the AMPK pathway
AMPK is a major regulator of not only glucose and lipid metabolism but also mitochondrial biogenesis [121314]. The AMPK pathway has been proposed to be the mechanism of the improvement in insulin resistance by Rg3 in skeletal muscle [615]. Therefore, we examined whether AMPK mediated the effect of Rg3 on mitochondria using compound C, an AMPK inhibitor. However, compound C did not alter the expression of mitochondrial complexes (Fig. 4), suggesting that Rg3 does not regulate mitochondrial biogenesis through the AMPK pathway.
DISCUSSION
In this study, we first demonstrated that Rg3 improves mitochondrial function and the expression of key genes involved in mitochondrial biogenesis in C2C12 myotubes. This might be the mechanism underlying Rg3-enhanced insulin signaling in skeletal muscle. Our findings posit a possible mechanism behind the improvement in insulin sensitivity by P. ginseng or ginsam: the regulatory effects of Rg3 on skeletal muscle mitochondria may mediate the anti-diabetic effects of P. ginseng or ginsam, at least in part.
Because the majority of glucose uptake occurs in skeletal muscle, insulin sensitivity in skeletal muscle is important to maintain plasma glucose levels [16]. Insulin binding to the insulin receptor leads to the phosphorylation of IRS-1, which subsequently activates the phosphoinositide 3-kinase/Akt pathway and results in increased glucose uptake [17]. We previously reported that Rg3 increased insulin sensitivity by inducing IRS-1 and Akt phosphorylation, and the subsequent expression of GLUT4 and glucose uptake in skeletal muscle [69]. In this study, we reconfirmed the activation of IRS-1 and Akt by Rg3 in C2C12 cells, and to elucidate the underlying mechanism of Rg3, we focused on the mitochondria.
Mitochondria play an important role in the insulin resistance of skeletal muscle [1011]. Reduced mitochondrial numbers and impaired mitochondrial function have been observed in the skeletal muscle of diabetes mellitus patients [11]. Therefore, we investigated changes in the mitochondrial biogenesis and function after Rg3 treatment. We found that Rg3 significantly increased ATP production and the OCR, indicating an enhancement of mitochondrial function in skeletal muscle. These results are consistent with those of a previous study, which reported that Rg3 enhanced ATP production and the respiratory control ratio in the rat brain [18].
Mitochondrial dysfunction can also impair fatty acid oxidation and increase the production of reactive oxygen species (ROS), leading to the accumulation of intramyocellular fat content and ROS, and consequently, insulin resistance. Previous studies have reported that Rg3 stimulates the phosphorylation of acetyl-CoA carboxylase, a key regulator of fatty acid oxidation in C2C12 myotubes [15] and cardiac muscle [19]. Rg3 also increased antioxidant enzymes in rat cardiac muscle [19] and decreased ROS production in brain mitochondria [18]. Although we did not examine fatty acid oxidation and ROS production, Rg3 is expected to improve fatty acid oxidation and ROS production based on the previous reports.
We also demonstrated that Rg3 improved the expression of key genes involved in mitochondrial biogenesis. In this study, Rg3 treatment increased the expression of PGC-1α and its downstream targets, NRF-1 and Tfam. PGC-1α is a key regulator that induces mitochondrial biogenesis by activating other transcription factors, such as NRF-1 and NRF-2, which subsequently activate the expression of Tfam [20]. Previous studies have demonstrated that Rg3 increases the expression of PGC-1α in cardiac muscle [19] and skeletal muscle [615]. Following the change in the expression levels of transcription factors, we observed an increase in both the mRNA expression and protein levels of some components of the mitochondrial complexes in C2C12 cells. While Rg3 increased the expression of key genes involved in mitochondrial biogenesis regardless of the pretreatment of antimycin A (Fig. 2), Rg3 improved ATP production or the OCR only under the antimycin A-treated condition (Fig. 3). There are several reasons why Rg3 improved mitochondrial function against antimycin A but not under the basal condition. One possibility is that stimulation of the mitochondrial biogenesis could not preserve or potentiate mitochondrial function under the basal condition but could facilitate the recovery of mitochondrial function after mitochondrial inhibition [2122]. Another possibility is the difference in the post-translational regulation according to the conditions.
AMPK senses the energy status of the cell through the AMP/ATP ratio in the muscle, and a low energy status, such as that produced during exercise, activates AMPK and regulates energy metabolism, including glucose uptake and mitochondrial biogenesis [2324]. AMPK is also known to stimulate PGC-1α through the cAMP-response element binding protein (CREB), subsequently increasing mitochondrial biogenesis [202526]. It is possible that AMPK mediates the effect of Rg3 on mitochondrial biogenesis. Rg3 or ginsam has been reported to activate AMPK phosphorylation in C2C12 myotubes [15] and the skeletal muscle of OLETF rats [6]. However, in our previous study [9], Rg3 did not activate AMPK, and compound C did not alter insulin signaling in L6 myotubes. Consistent with previous findings, AMPK seems not to mediate mitochondrial biogenesis in this study (Fig. 4).
In conclusion, we demonstrated that Rg3, a constituent of P. ginseng, improves mitochondrial biogenesis and function, leading to the improvement in insulin resistance in skeletal muscle. Therefore, Rg3 may contribute to the anti-diabetic effect of P. ginseng, and it may be a potential therapeutic agent for the treatment of type 2 diabetes mellitus.



 XML Download
XML Download