

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Fibroblast growth factor 21 (FGF21), along with FGF19 and FGF23, are endocrine factors that structurally belong to the FGF superfamily [1]. Unlike the classic FGFs that require heparin for stable binding to the FGF receptors (FGFRs), the three endocrine members of the FGF superfamily lack the heparin-binding property, and therefore can be released into the circulation [2]. These hormone-like FGFs are involved in the regulation of diverse metabolic pathways. FGF15/19 controls cholesterol/bile acid synthesis. FGF23 modulates phosphate/vitamin D metabolism, whereas FGF21 regulates glucose and lipid metabolism [2]. Physiologically, FGF21 expression is markedly increased in response to fasting/starvation, and elevated FGF21 is a central mediator for fasting-induced fatty acid oxidation, ketogenesis and for inducing growth hormone resistance [34]. Pharmacologically, therapeutic intervention with recombinant FGF21 counteracts obesity and its related metabolic disorders in both rodents and nonhuman primates, including reduction of adiposity and amelioration of hyperglycemia, hyperinsulinemia, insulin resistance, dyslipidemia, and fatty liver disease [5]. Furthermore, FGF21 is the downstream target of both peroxisome proliferator-activated receptor (PPAR) α and PPARγ, and a growing body of evidence suggests that the glucose-lowering and insulin-sensitizing effects of the PPARγ agonists (thiazolidinediones [TZDs]) and the therapeutic benefits of the PPARα agonists (fenofibrates) on lipid profiles are mediated in part by induction of FGF21 [67].
In addition to its multiple metabolic actions, emerging evidence suggests that FGF21 also possesses vascular protective activities, independent of its effects on glycemic control and insulin sensitivity. This review aims to summarize recent clinical and animal studies in this field, and discuss the molecular pathways underlying the antiatherosclerotic activities of FGF21.
BASIC BIOLOGY AND PHYSIOLOGY OF FGF21
FGF21 was initially identified as a metabolic regulator by a high throughput glucose uptake screen in 3T3-L1 adipocytes [8]. Although liver is a major source for circulating FGF21 [49], expression of FGF21 is also detectable in several other metabolic organs, including adipose tissue [1011], pancreatic islets [12], and skeletal muscle [13]. Due to lack of the heparin-binding property, FGF21 itself is not sufficient to bind the FGFRs, but requires the recruitment of the single-pass transmembrane glycoprotein βKlotho as a co-receptor [14151617], through its direction with the carboxyl terminus of FGF21 [18]. Both in vivo and in vitro studies have demonstrated the essential role of βKlotho in conferring the metabolic effects of FGF21 [14151617]. Unlike the ubiquitous expression pattern of FGFRs, βKlotho expression is restricted to a number of metabolic tissues (adipose tissue, liver, brain, and pancreas) [19], which may explain the tissue selectivity of the FGF21 targets.
The binding of FGF21 to its receptor complex has been shown to activate several downstream signaling pathways, including the mitogen-activated protein kinase (MAPK)/extracellular signal-regulated protein kinase 1/2 (ERK1/2) [8141520], protein kinase B (Akt) [12], adenosine monophosphate (AMP)-activated protein kinase, and sirtuin 1 (Sirt1) [21]. Although phosphorylation of ERK1/2 and its downstream targets has been widely used as a maker for FGF21 signaling, it is unlikely that ERK1/2 is a key player in mediating the multiple metabolic functions of FGF21. The molecular events that link the FGF21 receptor complex with its distal effects remain poorly characterized. It is also currently unclear why FGF21 does not have any mitogenic activity despite its ability to activate ERK1/2.
FGF21 AS A SENSOR OF METABOLIC STRESS AND A MEDIATOR OF MULTIORGAN CROSSTALK
In rodents, circulating levels of FGF21 are extremely low in fed status, but are markedly induced in multiple organs in response to diverse physiological or pathological stressors, such as starvation, nutrient excess, autophagy deficiency, mitochondrial dysfunction, exercise, drug toxicity, and cold exposure [22]. Starvation triggers lipolysis in adipose tissue to release free fatty acids, which in turn activate hepatic PPARα for production of FGF21. Elevated FGF21 released from the liver travels across the blood-brain barrier and enters the brain to activate hypothalamic neurons for production of corticotropin-releasing hormone, which in turn triggers the pituitary release of adrenocorticotropic hormone and subsequent secretion of corticosterone from adrenal gland. Corticosterone acts on the hepatocytes to promote hepatic gluconeogenesis [23]. Therefore, FGF21 maintain fasting glucose homeostasis by mediating the crosstalk between liver, adipose tissue and brain via activation of the hypothalamus-pituitary-adrenal (HPA) axis [2324].
In response to overdose of acetaminophen, both circulating levels of FGF21 and its hepatic production are drastically elevated [25], which in turn act in a compensatory manner to protect against the drug-induced hepatotoxicity by enhancing PGC-1α/Nrf2-mediated antioxidant capacity in the liver. Mitochondrial dysfunction caused by autophagy deficiency triggers the release of FGF21 from skeletal muscle, which in turn acts as a myokine to protect against high fat diet-induced obesity and insulin resistance possibly via its actions in adipose tissues [26]. On the other hand, cold exposure causes elevated expression in both brown and white adipose tissues, but not in the liver [27]. Notably, FGF21 has been identified as an obligatory mediator of environmental cold-induced browning and thermogenesis of white adipose tissues in mice [28]. However, it is currently unclear whether such a browning effect of FGF21 is attributed to its autocrine action adipose tissue, or endocrine actions via interorgan crosstalk. A recent study showed that the central actions of FGF21 may contribute to its browning activity in white adipose tissue [29], suggesting that FGF21 is also a mediator for the crosstalk between brain and adipose tissue.
FGF21 AS A PHYSIOLOGICAL PROTECTOR AGAINST ATHEROSCLEROSIS
Although early studies of FGF21 focused predominantly on its metabolic activities, emerging evidence suggests that FGF21 is also a physiological protector of vascular functions in both rodents and humans. In a study cohort including 670 Chinese subjects with measurement of carotid intima-media thickness, we have observed a positive correlation between elevated serum FGF21 levels and carotid atherosclerosis, independent of the traditional risk factors including smoking, creatinine, C-reactive protein, dysglycemia, and dyslipidemia [30]. Furthermore, a strong association of serum FGF21 levels with subclinical atherosclerosis and low-extremity atherosclerosis has also been observed in patients with type 2 diabetes [3132]. In line with these clinical observations, circulating levels of FGF21 are progressively elevated with ageing in apolipoprotein E-deficient mice (apoE-/-), a well-known model with spontaneous development of atherosclerosis [33].
To address the pathophysiological roles of FGF21 in the development of atherosclerosis, we have recently generated FGF21 and apoE double knockout (DKO) mice by crossing FGF21 knockout mice with apoE-/- mice in C57BL/J background [33]. Notably, both ageing- and western diet-induced atherosclerotic plaque formation (as determined by oil red O staining of entire aorta) in DKO mice is markedly exacerbated compared to that in apoE-/- littermates. Genetic depletion of FGF21 in apoE-/- mice also leads to premature death, possibly due to the development of atherosclerotic heart disease. In the atherosclerotic lesion areas of DKO mice, smooth muscle proliferation, macrophage infiltration, and collagen deposition are also significantly increased in comparison with apoE-/- mice [33]. There is no obvious difference in body weight, glucose tolerance and insulin sensitivity between DKO mice and apoE-/- mice on either standard or western diet. On the other hand, DKO mice exhibit markedly increased levels of proinflammatory chemokines (intercellular adhesion molecule-1 [ICAM-1], vascular cell adhesion protein-1 [VCAM-1], and monocyte chemotactic protein-1 [MCP-1]) and cytokines (tumor necrosis factor α [TNFα]) in both atherosclerotic lesion areas and bloodstream, and also worsened lipid profiles (hypercholesterolemia, decreased high density lipoprotein [HDL]/low density lipoprotein [LDL] ratio). All these changes can be reversed by treatment with recombinant FGF21. Taken together, these findings support the notion that FGF21 is a physiological protector against atherosclerosis possibly via its effects on alleviation of vascular inflammation and hypercholesterolemia, independent of adiposity, glycemic control, and insulin sensitivity. The elevated levels of circulating FGF21, as observed in both patients and animals with atherosclerosis, may serve as an adaptive mechanism to defend against this disease.
ADIPONECTIN AS A DOWNSTREAM EFFECTOR OF FGF21 IN PROTECTION AGAINST ATHEROSCLEROSIS
Despite the potent antiatherosclerotic activity of FGF21, its co-receptor βKlotho is hardly detectable in blood vessels (L.J. and A.X., unpublished data). Therefore, we tested the possibility that FGF21 may exert its vascular protective effects indirectly, via induction of adiponectin, which is one of the most abundant adipokines secreted from adipocytes. Circulating levels of adiponectin are decreased in obesity and a cluster of obesity-related cardiometabolic diseases [3435]. Epidemiological studies in different ethnic groups have reproducibly identified low levels of adiponectin (hypoadiponectinemia) as an independent risk predictor for type 2 diabetes, atherosclerosis and coronary heart disease [3436]. Conversely, elevation of circulating adiponectin by either genetic or pharmacological approaches can reverse insulin resistance, hyperglycemia, metabolic inflammation, atherosclerosis, myocardial infarction and stroke in either rodents or pig models [37]. In particular, adiponectin exerts its cardiovascular protect effects through its direct actions in almost all major types of cells in both the blood vessels and heart [3738]. In endothelial cells, adiponectin induces production of nitric oxide via activation of endothelial nitric oxide synthase [39], reduces high glucose-induced oxidative stress and suppresses expression of adhesion molecules and the cell interaction with leucocytes [40]. In macrophages, adiponectin suppresses both endotoxin- and toxic lipids-induced inflammation and blocks foam cell formation by inhibiting uptake of oxidized LDL and enhancing cholesterol efflux [41]. Furthermore, adiponectin inhibits growth factors-induced proliferation and migration of smooth muscle cells [42], and prevents platelet aggregation and activation. Notably, adiponectin-deficient mice are refractory to the protective effects of rosiglitazone against diabetes-induced vascular damage [43], suggesting an obligatory role of adiponectin in mediating the vascular protection of this PPARγ agonist.
Recent data from both us [44] and Scherer's group [45] has demonstrated the potent effects of FGF21 in inducing both expression and secretion of adiponectin. Notably, increased adiponectin is observed within 1 hour after administration with recombinant FGF21. Mechanistically, we found that both the acute and chronic effects of FGF21 on adiponectin production are mediated in part by PPARγ, a nuclear receptor that controls the gene transcription as well as the protein secretion of adiponectin [4647]. Furthermore, the stimulatory effects of the PPARγ agonists TZDs on adiponectin production in both mouse adipocytes and human adipose tissue are mediated by augmented autocrine actions of FGF21 [44]. In both fat explants isolated from PPARγ+/- mice and human fat explants pre-treated with the PPARγ antagonist GW9662, FGF21-induced production of adiponectin is partially abrogated, perhaps due to reduced expression of endoplasmic reticulum oxidoreductin-1-like α and disulfide-bond A oxidoreductase-like protein, both of which are molecular chaperones involved in adiponectin oligomerization and secretion [4849]. In support of this notion, a positive feedback loop between FGF21 and PPARγ in adipocytes has been shown to be critical in maintaining systemic glucose homeostasis [7]. The autocrine actions of FGF21 activate PPARγ possibly by modulation of sumoylation, which in turn leads to further induction of FGF21 expression via transcriptional activation. Consistent with these animal-based observations, a recent phase-1b clinical trial showed that treatment of obese human subjects with type 2 diabetes with LY2405319, a long-acting FGF21 analog, leads to a marked elevation in circulating adiponectin [50]. In adiponectin-deficient mice, both acute effects of FGF21 on decreasing hyperglycemia and hyperinsulinemia and chronic effects of FGF21 on alleviation of dietary obesity-induced insulin resistance, glucose intolerance, and fatty liver are severely impaired as compared to those in wild-type mice [44]. Similarly, the effects of FGF21 on reduction of ceramide and induction of energy expenditure are also dependent on adiponectin [45]. Taken together, these findings suggest that the metabolic benefits of FGF21 are mediated at least in part by induction of adiponectin.
In DKO mice, we found that the exacerbated atherosclerosis caused by FGF21 deficiency is significantly reduced by daily administration with either recombinant FGF21 or adiponectin [33]. However, the magnitude of reduction in atherosclerotic plaque size in adiponectin-treated mice is smaller than that in FGF21-treated mice. Further analysis showed that the effects of FGF21 and adiponectin administration on suppression of collagen composition, smooth muscle proliferation and macrophage infiltration, and reduction in expression of proinflammatory chemokines ICAM-1 and VCAM-1 and cytokines TNFα and monocyte MCP-1 are comparable. Consistently, our in vitro studies demonstrated that adiponectin, but not FGF21, exerts direct effects on suppressing proliferation and migration of human smooth muscle cells and uptake of oxidized LDL in human macrophages. On the other hand, while administration of recombinant FGF21 decreases total cholesterol in DKO mice to a level comparable to apoE-/- mice, whereas adiponectin has no effect on hypercholesterolemia caused by FGF21 deficiency. Therefore, these findings suggest that while adiponectin is critical for the protective effects of FGF21 on blood vessel cells, but is not required for the cholesterol-lowering activity of FGF21. Indeed, adiponectin does not have cholesterol-lowering activity despite its ability to reduce triglyceride [51].
LIVER AS A MAJOR TARGET OF FGF21 FOR SUPPRESSION OF HYPERCHOLESTEROLEMIA
Dyslipidemia, especially elevated LDL-cholesterol, is a major contributor to atherosclerotic plaque formation. The cholesterol-lowering drugs, such as statins, have been used clinically to reduce the risk of atherosclerotic heart disease. Therapeutic administration of FGF21 has been shown to alleviate dyslipidemia in rodents [52], obese monkeys [53], and obese patients with type 2 diabetes [50], including reductions in total and LDL-cholesterol and triglycerides, elevations in HDL-cholesterol and a shift to a less atherogenic apolipoprotein profile.
Consistent with the aforementioned pharmacological studies, FGF21 deficiency in apoE-/- mice causes a further aggravation of hypercholesterolemia and a shift of apolipoprotein profiles from HDL to LDL, which are accompanied by augmented de novo cholesterol biosynthesis and increased expression of several cholesterologenic genes in the liver, suggesting that endogenous FGF21 is a physiological suppressor of hepatic cholesterol production [33]. Mechanistically, we found that the inhibitory effects of FGF21 on hepatic cholesterol biosynthesis are attributed to its suppression of sterol regulatory element-binding protein 2 (Srebp-2), a member of the basic helix-loop-helix-leucine zipper transcription factor family and a master regulator of cholesterol biosynthesis by preferentially activating the transcription of key cholesterologenic genes [5455]. Adenovirus-mediated silencing of hepatic Srebp-2 expression is sufficient to counteract exacerbation of hypercholesterolemia and augmentation of hepatic cholesterol biosynthesis caused by FGF21 deficiency, whereas the therapeutic benefits of systemic FGF21 administration on inhibition of hepatic cholesterologenesis and reduction of hypercholesterolemia are abrogated by overexpression of Srebp-2. Interestingly, FGF21 specifically suppresses the transcription and activity of Srebp-2, but not Srebp-1, which preferentially activate transcription of genes involved in fatty acid synthesis [33].
Although FGF21 is predominantly produced from the liver, it is still debatable whether hepatocyte itself is a direct target of FGF21 [356]. While βKlotho is highly expressed in the liver, the expression of FGFR1, which is postulated as a major form of FGF21 receptor in adipocytes, is hardly detectable in this tissue [33]. Despite the fact that hepatic production of FGF21 is markedly induced upon prolonged fasting, FGF21 regulates hepatic gluconeogenesis indirectly via activation of the HPA axis, but not via its direct actions on hepatocytes [323]. On the other hand, direct effects of FGF21 on inhibition of gluconeogenesis and augmentation of fatty acid oxidation have also been reported [5758]. To address whether the regulatory effects of FGF21 on cholesterol homeostasis are attributed to its direct hepatic actions, we generated the βKlotho liver-specific knockout (βKlotho-LKO) mice by intravenous injection of adenovirus-associated virus encoding Cre recombinase into βKlotho-floxed mice [33]. Notably, the effects of FGF21 on reduction of hypercholesterolemia and inhibition of hepatic expression of Srebp-2 and several cholesterologenic genes are largely abrogated in βKlotho-LKO mice, despite a similar level of adiponectin between βKlotho-LKO and wild-type littermates. By using both gain-of-function and loss-of-function experiments, we identified FGFR2, which is highly expressed in hepatocytes, mediates the cholesterol-lowering effect of FGF21 in the liver. These findings collectively support the notion that the cholesterol-lowering effects of FGF21 are attributed to its direct actions on hepatocytes via the FGFR2-βKlotho receptor complex. However, the intracellular signaling pathways that link FGF21-mediated activation of the FGFR2-βKlotho receptor complex with the downstream Srebp-2 expression remain elusive.
CONCLUSIONS
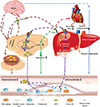
In addition to multiple metabolic benefits of FGF21, accumulating evidence from both animal and clinical studies demonstrate that this hormone also possesses potent protective effects against vascular inflammation and atherosclerotic lesions by mediating the crosstalk among several key metabolic and vascular organs (Fig. 1). First, FGF21 acts on its main target adipose tissues to promote the expression and release of adiponectin possibly by activation of PPARγ, and adiponectin in turn exerts its vascular protective activities via its direct effects on all the major types of cells in the blood vessels. Second, FGF21 alleviates hypercholesterolemia via its autocrine actions in hepatocytes, where it suppresses Srebp-2-mediated cholesterol biosynthesis via the FGFR2-βKlotho receptor complex. In addition, several other effects of FGF21 may also contribute indirectly to the antiatherosclerotic activity of FGF21, including its inhibition of lipolysis, which may in turn prevent lipotoxicity-induced metabolic and vascular damages [59]. Furthermore, FGF21 has been shown to promote the browning of white adipose tissues via either its direct actions in adipocytes or via mediating the crosstalk between brain and adipose tissues [272829]. Notably, transplantation of brown adipocytes has been shown to alleviate atherosclerosis in animals [60]. In humans, the amount and activity of brown adipocytes are inversely correlated with cardiovascular diseases [61].
Apart from the anti-atherosclerotic activities of FGF21, its cardiac protective effects have also been observed in several recent studies, including prevention of myocardial injury and apoptosis after ischemia reperfusion via FGFR1/βKlotho/Akt signaling cascade in cardiomyocytes [62]. FGF21 has also been reported to prevent cardiomyocytes apoptosis by activation of adiponectin-dependent signaling [63]. Furthermore, FGF21 alleviates diabetic cardiomyopathy by reducing cardiac lipid accumulation [64]. Taken together, these animal-based studies suggest FGF21 analogs represent a promising therapeutic agent for treatment and prevention of a cluster of cardiometabolic disorders. Although FGF21 was initially identified as glucose-lowering agent, a recent clinical in obese patients with type 2 diabetes showed that chronic administration of a long-acting form of FGF21 causes a marked elevation of adiponectin and an obvious reduction in total and LDL-cholesterol, but has little effect on hyperglycemia [50], further supporting the notion that FGF21 or its agonists might be more effective in reducing cardiovascular risks, instead of diabetes.
Despite the promising results on the cardiovascular protective effects of FGF21, it is important to note that almost all these studies are mainly based on rodent models. In light of the fact that there is a big difference in lipid metabolism and cardiovascular structure between rodents and humans, the pathophysiological relevance of these findings remains to be confirmed in humanoid large animals (such as pigs) and in clinical investigations. Furthermore, our knowledge on the receptor and post-receptor signaling events underlying the multiple physiological and/or pharmacological actions of FGF21 remain largely fragmented, and further extensive studies on FGF21 signaling cascade are needed to assist the rational design of FGF21 or its receptor agonists for future therapeutic applications to reduce the morbidity and mortality of cardiovascular diseases.
 XML Download
XML Download