

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Diabetic kidney disease (DKD) is the most common cause of end-stage renal disease worldwide [12]. It is characterized by a slowly increasing proteinuria and a gradual decrease of renal function. A number of factors important for the development of DKD, including hyperglycemia, hypertension, oxidative stress, and inflammation [3], have been shown to lead to proteinuria and histological changes, such as mesangial expansion, glomerular sclerosis, and tubulointerstitial fibrosis. Renal fibrosis is a final common pathologic finding observed in the later stage of disease and is characterized by transforming growth factor β (TGF-β)/Smad signaling-mediated extracellular matrix (ECM) accumulation [4]. Inhibition of the TGF-β/Smad signaling pathway is thus a promising therapeutic candidate for delaying the progression of DKD.
Dipeptidyl peptidase-4 (DPP-4) inhibitors increase the secretion of endogenous incretin, such as glucagon-like peptide-1 (GLP-1), which reduces post-meal blood glucose levels and elicits a variety of physiological actions. Recent studies demonstrated that the incretin system has protective effects against DKD. GLP-1 agonists exert renoprotective effects through anti-hypertensive [3], anti-inflammatory [56], and antioxidant actions [7]. DPP-4 is ubiquitous and is found in the kidney at relatively high concentrations [8910]. Because DPP-4 also cleaves substrates such as neuropeptide Y, high-mobility group protein B1, and meprin β, it may have pleiotropic effects [1112]. Several studies demonstrated the beneficial effects of DPP-4 inhibitors on various renal injuries. Treatment with DPP-4 inhibitors attenuated renal injury in DKD and improved renal fibrosis induced by either DKD or unilateral ureteral obstruction (UUO) [13141516]. A reduction in albuminuria has been observed following inhibition with DPP-4, which is a biochemical hallmark of DKD in patients with type 2 diabetes [1718]. Furthermore, a 6-month treatment of sitagliptin effectively ameliorated albuminuria in patients with type 2 diabetes compared with other glucose-lowering agents, despite no appreciable changes of glycemic control rate between the two groups [19]. In addition, alogliptin ameliorated albuminuria in patients with type 2 diabetes via up-regulation of stromal cell-derived factor-1α [20].
Gemigliptin is a potent, selective, competitive, and long-acting inhibitor of DPP-4 and is currently used for the treatment of type 2 diabetes [21]. It is administered as one daily dose, and its elimination is balanced between metabolism and excretion through urine and feces [22]. Regardless of its glucose-lowering effect, gemigliptin exerts pleiotropic effects on various diseases. Gemigliptin efficiently inhibited lipopolysaccharide-induced pro-inflammatory effects in vascular endothelial cells by attenuating nuclear factor (NF)-κB and JNK signaling via Akt/adenosine monophosphate-activated kinase dependent mechanisms [23] and prevented renal fibrosis in UUO models through inhibition of TGF-β and NF-κB activation [13]. However, there are very few studies on the effects of gemigliptin on a DKD model of type 1 diabetes. The aim of this study was to investigate whether gemigliptin has a renoprotective effect on DKD in a streptozotocin (STZ)-induced type 1 diabetic mouse model.
METHODS
Experimental design
Hyperglycemia was induced by a single intraperitoneal dose (150 mg/kg) of STZ (Sigma Aldrich, St. Louis, MO, USA) into 9-week-old male C57BL/6J mice (Japan SLC, Hamamatsu, Japan). Mice were orally administered 300 mg/kg/day gemigliptin (LG life science, Seoul, Korea) mixed with normal mice feedings for 8 weeks. Animals were split into three groups, each comprising six mice: the control group, STZ group, and STZ gemigliptin-treated group. Blood glucose levels and weight loss were monitored every 2 weeks. The mice were sacrificed using pentobarbital (50 mg/kg) at 8 weeks. Blood and kidney or pancreatic tissue samples were collected for histologic examination or frozen in liquid nitrogen for the isolation of protein or RNA. All procedures were performed in accordance with the institutional guidelines for animal research.
Measurement of renal function and biochemical markers
Blood glucose levels were measured 3 days following STZ administration using tail vein blood samples (Accu-Chek Active; Roche Diagnostics, Mannheim, Germany). Mice were housed in metabolic cages to collect urine for subsequent measurement of albumin or creatinine by enzyme-linked immunosorbent assay (ELISA). Creatinine in urine was determined using the Quantichrome Urea Assay Kit (BioAssay System, Hayward, CA, USA). Urine albumin kit was obtained from AssayPro (St. Charles, MO, USA). Plasma GLP-1 was measured using the appropriate ELISA kit in accordance with the manufacturer's instructions (LINCO Research, St. Charles, MO, USA).
Histologic and morphologic analysis
Kidneys were fixed with phosphate buffered saline containing 4% paraformaldehyde for overnight and then embedded in paraffin. Sections (4 µm) were cut and deparaffinized in xylene, followed by rehydration in a graded series of ethanol. Staining was performed using hematoxylin and eosin, periodic acid Schiff (PAS), and Sirius red staining. Immunohistochemical staining was performed as described previously [24], using type I collagen (Abcam, Cambridge, UK), fibronectin (BD Biosciences, San Jose, CA, USA), β-tubulin (Abcam), phospho-Smad3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), TGF-β (Santa Cruz Biotechnology), and insulin (Santa Cruz Biotechnology). Thickness of the glomerular basement membrane (GBM) was measured by electron microscopy (H-7100; Hitachi, Tokyo, Japan). Renal fibrotic areas were quantified by morphometric analysis using a light microscope equipped with an imaging system containing aMRc5 Carl Zeiss microscope (Oberkochen, Germany) and iSolution DT version 7.7 software (IMT i-Solution, Coquitlam, BC, Canada). Areas of positive PAS matrix, Sirius red, immunostaining for type I collagen, and fibronectin in the renal fibrotic regions (brown color) were quantified by computer-based morphometric analysis. All data were normalized to the control and expressed as fold increase relative to the control.
Cell culture
Rat mesangial cells (RMCs) and rat kidney proximal tubular epithelial NRK-52E cells were purchased from American Type Culture Collection (Manassas, VA, USA), and were cultured as described previously [24]. RMCs and NRK-52E cells were rendered quiescent by incubation for 24 hours in medium supplemented with 0.5% fetal bovine serum (FBS). Cells were incubated with the gemigliptin (100, 200, and 400 µg/mL) for 9 hours. Cells were treated with medium containing 0.5% FBS with or without TGF-β (5 ng/mL; Sigma) for 15 hours. Cells were subsequently processed for the isolation of RNA or protein as described below.
Quantitative real-time RT-PCR
Total RNA was isolated from RMC, NRK-52E cells, or mouse kidneys using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. cDNA was synthesized using a first-strand cDNA kit (Fermentas, Hanover, MD, USA). Quantitative real-time reverse transcription polymerase chain reaction (RT-PCR) was performed using the SYBR Green PCR Master Mix Kit (Applied Biosystems, Warrington, UK) on the StepOnePlus Real-Time PCR System (Applied Biosystems). The thermal cycling conditions were as follows: 95℃ for 10 minutes, followed by 40 cycles of 95℃ for 15 seconds and 60℃ for 1 minute. Primers were designed using AB StepOne software version 2.1 based on the relevant sequences from GenBank and were as follows: mouse type I collagen (sense: 5'-GCCTTGGAGGAAACTTTGCTT-3' and antisense: 5'-GCACGGAAACTCCAGCTGAT-3'; GenBank accession NM_007742.3), mouse fibronectin (sense: 5'-GATATCACCGCCAACTCATTCA-3' and antisense: 5'-CA GAATGCTCGGCGTGATG-3'; GenBank accession NM_ 010233.2), mouse glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (sense: 5'-GAAGGGTGGAGCCAAAAG-3' and antisense: 5'-GCTGACAATCTTGAGTGAGT-3'; GenBank accession NM_ 008084.2). Rat type I collagen (sense: 5'-GTGCGATGGCGTGCTATG-3' and antisense: 5'-TCGCCCTCCCGTTTTTG-3'; GenBank accession NM_ 053304.1), rat fibronectin (sense: 5'-ACCTGCAAGCCAATAGCTGAGA-3' and antisense: 5'-CCAGCCTTGGTAGGGCTTTT-3';GenBank accession NM_019143.2), rat GAPDH (sense: 5''-TGCCGCCTGGAGAAACC-3' and antisense: 5'-AGCCCAAGGA TGCCCTTTAGT-3'; GenBank accession NM_017008.4). The housekeeping gene GAPDH was used as an internal control.
Western blot analysis
For protein preparation, cells and kidney tissue were suspended in radioimmunoprecipitation assay buffer. The cells were then lysed on ice for 30 minutes, and the cell lysate was collected by centrifugation at 15,000 ×g for 10 minutes. Protein quantification was performed using a Bio-Rad Protein Assay kit (Bio-Rad, Richmond, CA, USA). Then, 30 µg of proteins were electrophoresed by sodium dodecyl sulphate-polyacrylamide gel electrophoresis and electrotransferred to polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). After blocking with 5% skimmed milk in Tris-buffered saline containing Tween 20 (0.1%) for 1 hour, the membrane was incubated with anti-type I collagen (1:1,000; Abcam), anti-fibronectin (1:1,000; BD Biosciences), anti-phospho-Smad3 (1:1,000; Cell Signaling Technology, Beverly, MA, USA), and anti-TGF-β (1:1,000; Cell Signaling Technology) polyclonal antibodies at 4℃ with gentle shaking overnight. Antibodies were detected by horseradish peroxidase-linked secondary antibody (Santa Cruz) using the enhanced chemiluminescence Western Blotting Detection System, in accordance to the manufacturer's instructions (Amersham, Buckinghamshire, UK). The membrane was reblotted with anti-β-tubulin antibody to verify equal loading of the protein in each lane. Densitometric measurements of the bands were made using the digitalized scientific program UN-SCAN-IT (Silk Scientific Corp., Orem, UT, USA).
RESULTS
Gemigliptin reduces albuminuria in STZ-induced type 1 diabetic mice
Treatment with gemigliptin (300 mg/kg) markedly increased serum GLP-1 in STZ-induced type 1 diabetic mice, suggesting that gemigliptin is capable of inhibiting DPP-4 activity (Fig. 1A). Blood glucose levels were significantly increased in mice 2 weeks after single injection of STZ (P<0.001 vs. control). Administration of gemigliptin did not reduce blood glucose levels (Fig. 1B). In addition, gemigliptin did not prevent pancreatic islet cell destruction induced by STZ (data not shown). Body weight of STZ-treated mice was decreased presumably due to marked hyperglycemia (Fig. 1C). Gemigliptin-treated STZ mice partially recovered body weight loss despite unimproved blood glucose levels (Fig. 1C). Gemigliptin-treated diabetic mice exhibited a marked decrease in urinary albumin and kidney weight compared with gemigliptin-untreated STZ-induced diabetic mice (Fig. 1D).
Gemigliptin treatment decreases GBM thickening
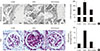
In STZ-induced diabetic mice, significant thickening of the GBM was observed (Fig. 2A). However, 8 weeks of treatment with gemigliptin prevented GBM thickening (Fig. 2A). Staining with PAS showed markedly attenuated mesangial ECM accumulation in kidney of gemigliptin-treated STZ mice compared with gemigliptin-untreated STZ mice (Fig. 2B).
Gemigliptin treatment attenuates STZ-induced renal fibrosis
To confirm the effects of gemigliptin on other DKD pathologic findings, we examined the effects of gemigliptin on STZ-induced renal fibrosis. H&E and Sirius red staining showed that gemigliptin treatment significantly reduced STZ-induced tubular atrophy and renal fibrosis (P<0.001) (Fig. 3A). Immunohistochemical staining demonstrated that gemigliptin-treated kidneys had reduced expression of type I collagen and fibronectin (Fig. 3B). The changes in mRNA and protein expression were further examined by real-time RT-PCR and Western blot analysis. Gemigliptin decreased STZ-induced type I collagen and fibronectin mRNA levels (Fig. 3C), and type I collagen and fibronectin protein expression (Fig. 3D).
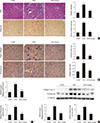
Gemigliptin decreases Smad3 phosphorylation in the kidney
To evaluate the mechanism by which gemigliptin attenuates renal fibrosis, TGF-β/Smad3 signaling was examined by immunohistochemical staining. An increase in TGF-β expression was observed in STZ-treated kidneys; however, no further ameliorations were observed in the kidneys from gemigliptin-treated diabetic mice (Fig. 4A). In contrast, renal expression of phosphorylated Smad3 (p-Smad3), a major downstream effector of TGF-β-mediated fibrosis, was significantly decreased in gemigliptin-treated diabetic mice (P<0.001) (Fig. 4A). To confirm these results, the changes were further examined by Western blot analysis. Gemigliptin had no effect on TGF-β protein expression, whereas p-Smad3 protein expression were significantly decreased (Fig. 4B).
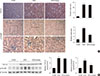
Gemigliptin inhibits TGF-β-stimulated type I collagen and fibronectin expression
We nextexamined whether gemigliptin inhibits TGF-β-stimulated Smad3 signaling in cultured renal cells. Gemigliptin inhibited TGF-β-stimulated mRNA expression and protein production of type I collagen and fibronectin in NRK-52E cells (Fig. 5A-C) and RMC cells (Fig. 5D-F) in a dose-dependent manner. Moreover, gemigliptin inhibited TGF-β-stimulated Smad3 phosphorylation in a dose-dependent manner in NRK-52E cells (Fig. 5B and C) and RMC cells (Fig. 5E and F). These results suggest that gemigliptin has a protective effect against renal fibrosis through the inhibition of TGF-β-stimulated Smad3 phosphorylation.
DISCUSSION
The study presented here evaluated the renoprotective role of gemigliptin in DKD regardless of its glucose-lowering effect. We found that gemigliptin treatment attenuates renal fibrosis in STZ-induced type 1 diabetic mice and was associated with a marked reduction of albuminuria, GBM thickening, and fibrosis. Furthermore, gemigliptin inhibited TGF-β-stimulated type I collagen and fibronectin mRNA expression and protein abundance and Smad3 phosphorylation in cultured renal cells, suggesting that gemigliptin protects against DKD via inhibition of the Smad3 pathway.
Apart from their essential role as glucose-lowering agents for type 2 diabetes, vildagliptin and linagliptin elicit their renoprotective effects independent of their glucose-lowering effect. STZ-induced diabetic rats, vildagliptin effectively inhibited early pathologic changes of DKD, such as GBM thickening, glomerulosclerosis, and interstitial expansion [15]. Linagliptin treatment in STZ-treated mice prevented the deterioration of renal structures and function [16]. In addition, linagliptin inhibited TGF-β-mediated endothelial-to-mesenchymal transition and renal fibrosis, indicating that linagliptin prevents late DKD in an insulin-deficient diabetes model in mice. In accordance with these findings, our present study showed that gemigliptin has a renoprotective effect on DKD regardless of a glucose-lowering effect in insulin-deficient diabetes. Gemigliptin reduced albuminuria and kidney weight in STZ-treated mice but did not reduce blood glucose levels, indicating that gemigliptin has beneficial effects on DKD independently of glycemic control. Furthermore, thickening of GBM was prevented in STZ-induced diabetes, implying that early administration of gemigliptin in insulin-deficient diabetes can prevent or retard early DKD. In addition, gemigliptin attenuated renal fibrosis and inhibited the in vivo expression of type I collagen and fibronectin in STZ-induced diabetes.
The TGF-β/Smad signaling pathway is a well-known mediator in the progression of DKD, mediating fibrosis by stimulating the synthesis of ECM molecules and by decreasing ECM degradation [25]. Although it has been identified that mesangial cells express Smad1, 2, 3, 4, and 7, recent evidence suggests that Smad3 is mainly implicated in pathogenic role in TGF-β-mediated renal fibrosis [42627]. A variety of molecular mediators and intracellular signaling pathways associated with DKD, such as hyperglycemia, angiotensin II, and oxidative stress, are connected and uniformly stimulate the TGF-β/Smad3 signaling pathway [25]. Consistent with the complexity and diversity of detailed molecular mechanisms by which these activities control TGF-β signaling, recent findings demonstrated that some DPP-4 inhibitors attenuate renal fibrosis via the inhibition of TGF-β expression levels in kidneys. Vildagliptin and linagliptin reduced TGF-β expression levels in STZ-induced diabetic mice kidneys [1516]. However, we showed that gemigliptin did not alter TGF-β expression in the kidneys of STZ-treated mice but reduced TGF-β-stimulated Smad3 phosphorylation and consequently decreased levels of ECM proteins, including type I collagen and fibronectin. The reason for a discrepancy of the TGF-β-lowering effect between DPP-4 inhibitors remains to be elucidated. In addition, given that TGF-β-induced renal fibrosis is well known to be closely associated with ECM protease matrix metalloproteinases, further studies are warranted to clarify the molecular mechanism responsible for gemigliptin's renoprotective effects on STZ-induced DKD.
This raises the question of whether the renoprotective effects of gemigliptin are GLP-1-mediated. Previous studies have shown an increase in local DPP-4 expression in the STZ-induced DKD, suggesting a negative pathological role [1516]. Although vildagliptin treatment increased plasma GLP-1 levels, opening the possibility of a GLP-1-dependent renoprotective mechanism [15], the latter study showed that linagliptin inhibited the TGF-β-induced endothelial-to-mesenchymal transition in a GLP-1-independent manner [16]. The renoprotective effects of gemigliptin in a UUO model have been demonstrated to be independent of GLP-1-mediated mechanisms [13]. It was suggested that the decreased expression of high mobility group box-1 by gemigliptin treatment was responsible for the protective mechanism. Because gemigliptin treatment was observed to increase plasma GLP-1 levels in the present study, we cannot exclude the possibility of the DPP-4 inhibitory effect of gemigliptin in the kidney. This issue may be resolved by adopting a tissue-specific DPP-4 deficient model.
In conclusion, our data showed that gemigliptin has a renoprotective effect on DKD through inhibition of Smad3, regardless of its glucose-lowering effect. These results suggest that gemigliptin could provide a therapeutic role in the prevention of DKD, including in patients with type 1 diabetes.


 XML Download
XML Download