

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Diabetic retinopathy (DR) is a leading cause of vision loss in working-age populations. In patients with diabetes, the overall prevalence of DR is approximately 35% [1]. While DR is clinically categorized into nonproliferative and proliferative types, diabetic macular edema (DME; present in 25% of persons with diabetes) affects central vision with metamorphopsia and micropsia in any type of DR [2]. In proliferative DR, obstruction of retinal capillaries leads to upregulation of proangiogenic growth factors, including vascular endothelial growth factor (VEGF), in hypoxic retinas [3]. Consequently, neoangiogenesis, an outgrowth of new blood vessels into the posterior surface of the vitreous, directly causes vision-threatening hemorrhage and retinal detachment [4]. While laser photocoagulation has been the standard modality to suppress DME and neoangiogenesis, intraocular injections of anti-VEGF drugs, either alone or in combination with surgical interventions, have globally become an additional option [5,6]. Although the intraocular anti-VEGF therapy has raised awareness of the feasibility of pharmacological treatment for DR, its limited efficacy has evoked the necessity to identify new drug targets for the fundamental treatment of DME and neoangiogenesis [7]. While diabetes animal models fail to reproduce human DR pathophysiology, technical advances in molecular manipulations in postnatal mouse retinas have enabled recapitulation of retinal edema and neoangiogenesis without hyperglycemia. This article will review the experimental model systems utilizing mouse retinas and discuss perspectives on new drug discovery for the treatment of DR.
ANGIOPOIETINS AS DRUG TARGETS FOR RETINAL EDEMA
Pericyte dropout in retinal vessels
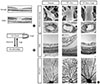
In DR, impairment of retinal vascular integrity causes elevated vascular permeability, which subsequently leads to fluid accumulation in extracellular spaces and swelling of retinal neurons and glial cells in the macula (Fig. 1A). Based on the postmortem examination of patients with DR, it was proposed that dropout of periendothelial mural cells (pericytes) from capillary walls might be an initial pathological event followed by deterioration of vessel integrity [8]. However, while DR clinically becomes manifest more than 5 years after the onset of diabetes, experimentally hyperglycemic rats or mice, with a 2-year average life expectancy, fail to represent human DR. Therefore, the widely-accepted 'pericyte dropout' concept had not been experimentally confirmed in diabetic animal models.
Retinal vascular changes caused by the absence of pericytes
To overcome the limitation of diabetic animal models, we assessed the retinal vascular changes primarily caused by pericyte dropout in postnatal mouse retinas, in which new blood vessels start to grow just after birth [9,10]. In newly-formed retinal vessels, vascular endothelial cells (ECs) secrete platelet-derived growth factor (PDGF)-B to facilitate the proliferation and migration of pericytes that express PDGF receptor β (PDGFRβ) (Fig. 1B) [11-13]. While Pdgfb knockout mice demonstrate defective pericyte recruitment in brain capillaries [14], the perinatal lethality of mice genetically deficient for Pdgfb or Pdgfrb precluded observations of postnatal vascular development in retinas [15,16]. Thus anti-PDGFRβ monoclonal antibodies were administered daily intraperitoneally to neonatal mice. This pharmacological blockade of PDGFRβ signal did not affect the overall growth of the neonates, but successfully disrupted pericyte recruitment to developing retinal vessels (Fig. 1C) [17]. Remarkably, the retinal vasculature devoid of pericytes recapitulated most of the pathophysiological features characteristic of DR; i.e., disorganized vascular networks with vessel dilation and tortuosity, and progressive extravasations with retinal edema and hemorrhage. Electron microscopic observations of pericyte-free retinal vessels further depicted thickening of periendothelial basement membranes, which was also seen in human DR. These data, obtained by pharmacological PDGFRβ manipulations, together with EC-specific ablation of the Pdgfb gene [18], indicated that pericyte dropout was sufficient to reproduce retinal vascular abnormalities in DR, even without hyperglycemia.
Ang1 restores the integrity of pericyte-free retinal vessels
In nascent vascular walls, pericytes contribute to stabilizing endothelial integrity via soluble signaling molecules and direct cell-cell contacts. In particular, angiopoietin-1 (Ang1) derived from pericytes binds to Tie2 receptor tyrosine kinase on EC surfaces, thereby activating downstream signals required for EC stabilization (Fig. 1B) [19,20]. Given that the absence of pericytes eliminates all of the pericyte-derived signals, we assessed to what extent Ang1 supplementation could restore the retinal vascular abnormalities caused by pericyte dropout. To our surprise, intraocular injections of recombinant Ang1 protein in conjunction with systemic injections of anti-PDGFRβ antibodies resulted in dramatic restoration of an organized architecture of retinal vascular networks [17]. Moreover, retinal edema and hemorrhage were completely suppressed despite the absence of pericytes (Fig. 1C). These experimental results indicated that Ang1 alone can maintain the structural integrity of retinal vessels in the complete absence of pericytes, and further suggested the potential efficacy of intraocular Ang1 therapy for the treatment of DME.
Ang2 as a potential target for the treatment of DR
Although PDGF-B/PDGFRβ signal is a prerequisite for pericyte recruitment to growing retinal vessels, a PDGFRβ blockade failed to deplete pericytes in adult retinas. Thus, alternative signals other than PDGF-B/PDGFRβ might be involved in the maintenance or disruption of EC-pericyte association in DR. In the adult vasculature, Ang1 derived from pericytes constitutively phosphorylates Tie2 at a low level to maintain the mature phenotype of the endothelium [21]. In contrast to the stable Ang1 expression in quiescent vessels, Ang2, a natural antagonist of Ang1, is expressed predominantly in ECs of activated blood vessels (Fig. 1B) [22-24]. Because Ang2 binding does not activate Tie2 in ECs, it was proposed that Ang2 destabilizes EC-pericyte association by interfering with Ang1, thereby rendering ECs highly sensitive to the microenvironment [22]. Specifically, Ang2 promotes neoangiogenesis and vascular leakage in the presence of VEGF and proinflammatory cytokines, but facilitates vascular regression in the absence of VEGF [25]. Importantly, while Ang2 expression is upregulated by hypoxia and VEGF [26,27], high glucose directly upregulates Ang2 transcription in cultured ECs [28]. Thus, in diabetic patients, it is plausible that hyperglycemia might induce Ang2 expression in retinal ECs, thereby destabilizing the ECMC association. In this scenario, a pharmacological Ang2 blockade would be of therapeutic benefit for the prevention of pericyte dropout in DR. To date, several pharmaceutical companies have developed anti-Ang2 drugs, some of which are under clinical trial for the treatment of tumor angiogenesis [29]. In the near future, it is expected that the therapeutic potency of anti-Ang2 drugs will be clinically evaluated for the treatment of DR.
NEOANGIOGENESIS IN ISCHEMIC RETINAS
Extraretinal neoangiogenesis in DR
In accordance with the progression of DR, retinal capillaries are obstructed, generating nonperfused, ischemic retinal areas. In response to hypoxia, retinal neurons, and glial cells secrete a series of proangiogenic growth factors, including VEGF, which leads to the formation of new blood vessels from pre-existing ones. However, these new vessels do not grow into ischemic retinas, but grow out of the retinal surfaces, without resolving retinal hypoxia. Moreover, the extraretinal vessels directly cause vitreous hemorrhage and tractional retinal detachment, both of which severely impair vision. Therefore, for the prevention and regression of neoangiogenesis, laser photocoagulation is performed in an attempt to destroy hypoxic neurons thereby reducing oxygen demands [30]. More recently, intraocular anti-VEGF drugs have been administered prior to vitreous surgery to lessen the intraoperative and postoperative hemorrhagic complications [31]. Nevertheless, the ideal way to fundamentally resolve retinal ischemia would be to rectify the angiogenic directions into the hypoxic retinas, thereby regenerating functional capillaries. To develop such a vascular regeneration therapy, useful insight can be obtained from the procedures of intraretinal angiogenesis during development.
Intraretinal angiogenesis during development
In the developing retinal vasculature, the angiogenic directions are determined by the endothelial filipodia projecting from the tips of sprouting blood vessels [11]. The tip ECs continuously project and retract numerous filopodia in response to the microenvironmental cues (Fig. 2A). The predominant force of endothelial filopodia projections is the concentration gradient of VEGF along the pre-existing networks of astrocytes, which migrate into the retina from the optic nerve prior to the onset of retinal vascular development [32]. In addition to VEGF secretion, astrocytes facilitate intraretinal angiogenesis by depositing extracellular matrices that serve as physiological scaffolds for migrating ECs [33]. As a consequence, developing vessels grow into hypoxic retinas in an organized fashion utilizing the astrocyte network as a template. Collectively, intraretinal angiogenesis largely depends on the signaling cues that regulate endothelial filopodia projections and the formation of matrix scaffolds by astrocytes (Fig. 2B).
Proangiogenic properties of retinal astrocytes
During development, retinal astrocytes act as proangiogenic cells in response to hypoxia, which is corroborated by previous observations that retinal astrocytes are found only in species that develop vascularized retinas, such as humans and rodents [34,35], whereas deficits in astrocytes resulted in the failure of retinal vascular development in postnatal mice [36]. In unvascularized areas of developing retinas, astrocytes express VEGF and fibronectin, a major component of matrix scaffolds for migrating ECs that express α5β1 integrin. However, expression levels of these proangiogenic genes in retinal astrocytes are rapidly downregulated upon contact with growing blood vessels. This indicates that oxygenation or extrinsic signals derived from blood vessels might deprive retinal astrocytes of proangiogenic activities. Notably, in postnatal mouse retinas, an orphan nuclear receptor Tlx is expressed in proangiogenic astrocytes of unvascularized, but not vascularized, areas [37]. Furthermore, the expression levels of the Tlx and Vegf genes in astrocytes were synchronously controlled by the oxygen concentration in postnatal mouse retinas. In Tlx-deficient mice, gene expression, as well as the extracellular protein transport of fibronectin, was severely impaired in retinal astrocytes, which led to defects in intraretinal vascular growth [37]. These results together indicated that Tlx is a hypoxia-inducible proangiogenic switch that controls the formation of extracellular fibronectin scaffolds in retinal astrocytes (Fig. 2C).
Signaling cues regulating endothelial filopodia formation
In developing retinas, VEGF isoforms with positively-charged domains encoded by exons 6 and 7 bind to the heparan sulfate proteoglycans, thereby constructing concentration gradients along the astrocyte network [11,38]. On the other hand, soluble VEGF isoforms lacking the heparin-binding domain disturb VEGF concentration gradients. Indeed, overexpression of soluble VEGF isoforms leads to disoriented projections of endothelial filopodia [11,38]. To restrict the pathfindings of retinal vascular growth, a member of axon guidance molecules, including slit, netrin, and semaphorin, have been implicated in the retraction of disoriented endothelial filopodia [39]. Especially, a soluble Sema3E ligand, derived from the retinal neurons, mediates filopodia retraction by binding to the endothelial PlexinD1 receptor [40-42]. Because PlexinD1 expression is upregulated by VEGF in ECs of developing retinal vessels [40,41], it is postulated that Sema3E-PlexinD1 signals form a negative feedback loop against VEGF-induced filopodia projections in retinal ECs.
Intracellular signaling machinery regulating endothelial filopodia formation
Projections and retractions of endothelial filopodia involve rearrangements of actin cytoskeletons and cell-matrix adhesions, which are regulated by Rho family small GTPases [43]. While Cdc42 promotes filopodia projections downstream of VEGF signals [44], RhoJ mediates endothelial filopodia retraction downstream of Sema3E signals [40]. Intriguingly, the activation states of Cdc42 and RhoJ were inversely regulated by VEGF and Sema3E (Fig. 2D). Because both Cdc42 and RhoJ bind to the Cdc42/Rac interactive binding domain of p21-activated kinases and neural Wiskott-Aldrich syndrome protein [45,46], it seems likely that Cdc42 and RhoJ compete for binding to their common downstream effector molecules, thereby controlling the dynamic rearrangements of actin cytoskeletons and cell-matrix adhesions. Given that Rho family small GTPases are activated by guanine nucleotide exchange factors (GEFs) [47], small molecule inhibitors targeting endothelial GEFs are expected to have therapeutic value for the control of endothelial filopodia projections. While a number of endothelial RhoGEFs have been identified [48], Arhgef15 acts as an EC-specific GEF to mediate VEGF-induced Cdc42 activation and further potentiates RhoJ inactivation, thereby promoting actin polymerization [49]. Because inactivation of the Arhgef15 gene resulted in retardation of retinal vascular growth [49], Arhgef15 would be a potential drug target for the treatment of aberrant neoangiogenesis in DR.
Regeneration of functional capillaries in the ischemic retina
In an oxygen-induced retinopathy (OIR) model that represents neoangiogenesis in ischemic retinas [50], intraocular anti-VEGF drugs suppress not only the extraretinal neoangiogenesis but also the desirable regeneration of intraretinal vessels, resulting in the deterioration of retinal ischemia [40]. These experimental results raise caution for the clinical use of anti-VEGF drugs in the treatment of proliferative DR. In the OIR model, while Sema3E was expressed in retinal neurons, PlexinD1 and RhoJ were highly expressed in ECs of extraretinal vessels [40]. Because extraretinal neoangiogenesis was enhanced by inducible EC-specific Plxnd1 loss of function but was diminished by inducible EC-specific Rhoj gain of function [40], it seems likely that binding of neuron-derived Sema3E to endothelial PlexinD1 activates RhoJ, thereby suppressing extraretinal vascular outgrowth. By exploiting PlexinD1 in extraretinal vessels as a molecular target, it was further demonstrated that intraocular injections of Sema3E proteins suppressed aberrant neoangiogenesis without affecting intraretinal regeneration of functional capillaries (Fig. 2E) [40]. These results implied a clinical application of intraocular Sema3E therapy for the treatment of proliferative DR.
CONCLUSIONS
Experimental manipulations in developmental and ischemic retinal models in mice have provided invaluable information to understand the pathophysiology of retinal edema and neoangiogenesis. However, there remain a number of gaps to be filled between these animal models and human DR. For instance, the involvement of hyperglycemia in retinal vascular dysfunctions is yet to be elucidated. In the future, the combination of diabetic animal models with molecular manipulations will give further insights into the molecular and cellular events underlying the onset and progression of DR, by which new drug targets will successfully be determined.

 XML Download
XML Download