

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In both developing and developed countries, calorie consumption exceeds the necessary level. In addition, modern sedentary lifestyle predisposes people to obesity. The spread of obesity is strongly associated with increasing prevalence of type 2 diabetes mellitus (T2DM) [1,2]. Obesity induces insulin resistance and thus insulin hypersecretion in pancreatic β-cells, resulting in β-cell dysfunction, particularly in people who are genetically susceptible to T2DM [3-6]. Therefore, controlling obesity may be critical in preventing an increase in the number of T2DM patients.
Green tea extract (GTE) has been widely considered as a potential supplemental remedy for obesity and T2DM; however, its effectiveness in humans has not been clearly demonstrated. In a previous study, we found that GTE reduces postprandial blood glucose levels (BGLs) in the intestinal lumen but elevates postprandial BGLs in the circulation [7]. In addition, we found that gallated catechins (GCs) in GTE, such as epigallocatechin gallate (EGCG) and epicatechin gallate (ECG), could be the major active components [7]. GCs inhibit the function of plasmalemmal glucose transporters in most cells, including Na-dependent glucose transporters in the intestine and kidney (where the GCs exert their beneficial effect), and Na-independent glucose transporters (GLUTs) in the liver, skeletal muscle and adipocytes, where GCs block the cellular entry of glucose [8]. Therefore, nontoxic polymers may be needed to block the intestinal absorption of GCs, lessening the inhibitory effect of GCs on various GLUTs in the circulation.
Previously, it was found that the antibacterial activity of GTE against Helicobacter pylori observed in vitro was not observed in vivo, most likely because GTE does not stay long enough in the gastrointestinal lumen, instead being easily absorbed into the circulation [9]. When sucralfate (sucrose sulfate-aluminum complex), which is used for antiulcerative therapy and has an affinity for the mucosal membrane of the gastrointestinal tract, is coadministered with GTE, the antibacterial activity of GTE can be observed in vivo [10]. This novel finding prompted us to use poly-γ-glutamic acid (γ-PGA) to increase the persistence of GCs in the intestine and inhibit the intestinal absorption of GCs. γ-PGA is a natural component of food and has been proven to be safe for animals and humans. It is a mucin-like amino acid chain in which D- and L-glutamic acid are linked by a γ-glutamyl bond. γ-PGA can be obtained by cultivating Bacillus subtilis from foods made with fermented beans (Japanese natto, Nepali kinema, Korean chungkookjang for B. subtilis var. chungkookjang). γ-PGA has been widely used as a supplement in foods, cosmetics and drug-delivery systems [11-13]. In this study, we aimed to evaluate the effect of a 4-week GTE+γ-PGA regimen on gain in body weight (BW) and glucose intolerance in obese T2DM db/db mice compared to treatment with GTE alone.
METHODS
Chemicals and diets
γ-PGA (MW: 2,000 KDa) was delivered from NUC Electronics Co. (Daegu, Korea). Green tea leaves (Bosung Seijak) were purchased from Bosung Green Tea Co. (Bosung, Korea). A normal commercial chow diet (NC; 10% fat, 70% carbohydrate, 20% protein), a normal chow diet containing γ-PGA (1 g γ-PGA/kg diet), a normal chow diet containing GTE (10 g GTE/kg diet), and a normal chow diet containing GTE+γ-PGA (10 g GTE/kg and 1 g γ-PGA/kg diet) were prepared by Hyochang Science (Seoul, Korea). Briefly, each ingredient was weighed and blended to homogeneity into appropriately powdered diet. The mixture was then formed into equal sized pellets and placed into a temperature- and humidity-controlled room to remove moisture.
Preparation of GTE
For animal studies, 20 g of green tea leaves was added to 1,000 mL of nanopure water. The tea leaves were stirred for 5 minutes at 80℃ and removed by filtration using filter paper (Advantec 2 filter paper; Hyundai Micro Co., Seoul, Korea) under reduced pressure. The extract was dried by lysophilization. A total of 3 g of GTE was harvested. The procedure was repeated until enough amount of GTE necessary for the experiments were harvested; the levels of EGCG, ECG, epigallocatechin, and epicatechin were 100, 53, 56, and 31 mg/g GTE, respectively. Periodically, the diet containing GTE was checked to confirm the concentrations of catechins exceeding 200 mg/g GTE.
Analysis of catechins from GTE with high performance liquid chromatography
High performance liquid chromatography (HPLC) analysis was conducted on a Waters Alliance 2,695 liquid chromatograph equipped with a model 2,487 dual absorbance detector (Waters Co., Milford, MA, USA). A Waters Symmetry C18 reversed-phase packing column (4.5×250 mm, 5 m) was used at 25℃ for separation throughout this study. Catechins were determined simultaneously at 235 nm. A gradient elution was performed by varying the proportion of solvent A (water-trifluoroacetic acid, 99.9:0.1 v/v) to solvent B (acetonitrile-trifluoroacetic acid, 99.9:0.1 v/v), with a flow rate of 1 mL/min. The mobile phase composition changed linearly from 9.5% to 14% solvent B in 10 minutes and then kept the same composition for 10 minutes. There was a linear increase in the proportion of solvent B, which reached 27.5% within 15 minutes. The mobile phase composition then returned to the initial conditions over a period of 5 minutes for the next run. All prepared solutions were filtered through 0.45 µm membranes (Sartorius, Maisemore, UK), and the mobile phase was degassed before injection into the HPLC.
Nuclear magnetic resonance spectroscopy of EGCG-γ-PGA complex
Nuclear magnetic resonance (NMR) spectroscopy was used to investigate the molecular interactions between EGCG and γ-PGA. 1H NMR and 13C NMR spectra were measured on a JEOL JNM-AL 300 (300 MHz) spectrometer. The sample for EGCG-γ-PGA (w/w, 1:1) complex was prepared by dissolving it in D2O/DMSO-d6 (1:1; Sigma-Aldrich, St. Louis, MO, USA) (concentration of 0.25% [w/v]).
Animals and treatments
To determine the short-term effects of GTE+γ-PGA on type 2 diabetic mice, C57BLKS/J Leprdb/Leprdb mice (db/db, male, 9 weeks old, 30.0 to 35.0 g, BGLs 200 to 300 mg/dL) and age-matched control nondiabetic heterozygous mice (male, 18.0 to 22.0 g, BGLs 110 to 140 mg/dL) were purchased from Jung-Ang Experimental Animals (Seoul, Korea) and used as an obese diabetic mouse model. The mice were allowed to acclimate for 1 week on chow and water. Starting at 10 weeks of age, the mice were randomly divided into four groups of 10: control, γ-PGA, GTE, and GTE+γ-PGA. The mice were provided with a semisynthetic normal chow diet containing vehicle, γ-PGA, GTE, or GTE+γ-PGA for 4 weeks. The food consumption of individual mice was checked every day. BW was measured every 7 days using an electronic balance. All mice had free access to food and water and received their specified diet for 4 weeks. The animals were housed (3 to 4 per cage) under a daily cycle of 12 hours light and 12 hours darkness. The experiment conditions were approved by the Keimyung University Institutional Ethics Committee, Daegu, Korea, which supervises animal research.
Oral and intraperitoneal glucose tolerance test
Oral glucose tolerance test (OGTT) was performed on normal mice. The mice were randomly divided into six groups of seven each: control, GTE, GTE+γ-PGA, GTE+1/2γ-PGA, GTE+1/3γ-PGA, and GTE+1/4γ-PGA. On the test days, GTE (900 mg/kg containing 90 mg EGCG/kg) and γ-PGA (90, 45, 22.5, and 11.25 mg/kg) were suspended in water and administered orally to fasted (12 hours) mice. Ninety minutes later, 2 g/kg glucose was given orally. The BGLs were measured in tail blood samples collected at 0, 15, 30, 60, 90, 120, and 180 minutes after glucose treatment. BGLs were measured using Glucocard Test Strip II (Arkray Inc., Kyoto, Japan). Intraperitoneal glucose tolerance test (IPGTT) was performed after the 4-week drug treatment in nondiabetic control and db/db mice. On test days, the animals were fasted for 12 hours and then given an intraperitoneal injection of glucose (500 mg/kg). BGLs were measured in tail blood samples at 0, 15, 30, 60, 90, 120, and 180 minutes after the glucose treatment.
Collection of blood and internal organ samples
At the conclusion of the study, animals were anaesthetized (nembutal, 100 mg/kg), and a 500-µL blood sample was drawn from the orbital venous plexus into a heparinized tube and immediately chilled on ice. The plasma was then separated and stored at -80℃ before analysis. After blood sampling, visceral adipose tissues (perirenal, retroperitoneal, and epididymal depots), liver, pancreas, and hypothalamus were dissected. The hypothalamus tissues were immediately frozen in liquid nitrogen and stored at -80℃ until measurement of mRNA levels by real-time reverse transcriptase-polymerase chain reaction (RT-PCR). Visceral adipose tissues were weighed and immediately frozen in liquid nitrogen. The liver and pancreas were fixed immediately in 10% neutral formalin solution and then embedded in paraffin.
Biochemical profile
Serum samples were obtained from blood by centrifugation for 15 minutes at 950 g at room temperature. Fasting glucose and insulin were measured using commercial kits (SpinReact, Gerona, Spain; Millipore, Billerica, MA, USA, respectively). The homeostasis model assessment of insulin resistance (HOMA-IR) was calculated using the final blood glucose and insulin levels in food-deprived mice.
Real-time polymerase chain reaction analysis
Each whole mouse hypothalamus was homogenized in TRI reagent (Sigma-Aldrich) using Ultra-Turrax T25 (IKA, Staufen, Germany). RNA was reverse transcribed to cDNA from 1 µg of total RNA using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). Quantitative real-time PCR was performed using Real-Time PCR 7500 System and Power SYBR Green PCR Master Mix (Applied Biosystems) according to the manufacturer's instructions. The expression level of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. The reactions were incubated at 95℃ for 10 minutes, followed by 45 cycles of 95℃ for 15 seconds, 55℃ for 20 seconds, and 72℃ for 35 seconds. Primers for mouse neuropeptide Y (NPY) and GAPDH were based on NCBI's nucleotide database and designed using the Primer Express program (Applied Biosystems): mouse NPY (forward, 5'-TAT CCC TGC TCG TGT GTT TG-3'; reverse, 5'-GTT CTG GGG GCA TTT TCT G-3'), mouse GAPDH (forward, 5'-CAG GAT GAC ACC AAA ACC CTC-3'; reverse, 5'-TCC AAG CCA GTG ACC CTC TG-3').
Liver and pancreas histopathology
Embedded liver and pancreas tissue blocks were cut into 6-micron sections and stained with hematoxylin and eosin. Pancreas tissue blocks were stained with HRP conjugated anti-insulin antibody. Mounting medium and cover slips were placed on the slides, which were then left to dry overnight. A diagnosis of fatty liver was made based on the presence of macrovesicular or microvesicular fat in 0.5% of the hepatocytes in a given slide.
Tissue lipid determination
Samples of liver were homogenized in 0.25% sucrose containing 1 mM ethylenediaminetetraacetic acid. Lipids were extracted using chloroform/methanol methanol (2:1, v/v) and evaporated in Savant Speedvac Concentrator (Thermo Fisher Scientific Co., Waltham, MA, USA). Next, the pellets were dissolved in 5% fatty acid-free bovine serum albumin dissolved in water. Proteins in the homogenate were assayed using protein assay reagent (Bio-Rad, Hercules, CA, USA) to normalize the amount of lipid extracted. Tissue triglyceride levels were determined using kits from Roche Diagnostics.
Statistical analysis
The results are expressed as the mean±standard error. SPSS version 14.0 (SPSS Inc., Chicago, IL, USA) was used for statistical analyses. The area under the curve (AUC) was calculated using Microcal Origin software version 7.0 (Microcal Software, Northampton, MA, USA). Comparisons between the two groups were performed with Student two-tailed t-test. When comparing two or more groups, significance was tested using ANOVA with Bonferroni correction to deal with relatively small number of samples. P values of less than 0.05 were considered significant.
RESULTS
Effect of GTE+γ-PGA on OGTT in normal mice
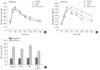
When glucose was loaded orally 90 minutes after each drug treatment, BGL significantly increased in GTE group compared to control (Fig. 1A and B), suggesting that GCs are already absorbed into the circulation where they inhibit cellular glucose uptake in various tissues. However, BGL of the GTE+γ-PGA groups were lower than the control group in γ-PGA-dose-dependent manner, suggesting that γ-PGA captures GCs in the alimentary tract, inhibiting glucose absorption. NMR measurement revealed possible interaction sites between γ-PGA and EGCG molecules through hydrogen bond and van der Waals interaction (Fig. 1C).
Effect of GTE+γ-PGA on food intake, BW, and body fat in db/db mice
There were no differences in BW, fed blood glucose, and daily food intake among the groups at the beginning of the experiment (data not shown). Obesity and diabetes were verified by evaluating BW and BGLs, respectively. The db/db mice with BGLs of 250 mg/dL or greater were considered as diabetic. After the 4-week drug treatment, the GTE+γ-PGA diet db/db mice showed approximately 15% less BW gain compared to control diet db/db mice (Fig. 2A and B). This was attributable to weight decreases in visceral adipose tissues containing perirenal, retroperitoneal and epididymal depots (Fig. 2C and D). However, GTE alone did not have any effect on BW or visceral fat contents. Daily food intake per cage did not differ among the groups (Table 1). Hypothalamic NPY, a typical orexigenic peptide [14,15], was also analyzed-there was no significant difference in basal hypothalamic NPY expression between the four groups of db/db mice after the drug treatment (Table 1).
Effect of GTE+γ-PGA on fasting blood glucose, fasting plasma insulin, and HOMA-IR index
After 4 weeks of the drug treatments, overnight fasting blood glucose and insulin levels were significantly lower in the GTE+γ-PGA diet db/db mice compared to control diet db/db mice (Fig. 3A and B). Additionally, HOMA-IR, a marker of insulin resistance, was also lower in GTE+γ-PGA diet db/db mice compared to control diet db/db mice (Fig. 3C).
Effect of GTE+γ-PGA on glucose tolerance
Glucose tolerance in nondiabetic control (Fig. 4A) and db/db (Fig. 4B) mice was evaluated by IPGTT after 4 weeks of treatment. GTE+γ-PGA diet in db/db mice significantly ameliorated glucose intolerance compared to the control diet (Fig. 4B). The AUC describing the blood glucose of the GTE+γ-PGA db/db group was approximately 25% less (P<0.01) than that of the control group (Fig. 4C).
Effect of GTE+γ-PGA on pancreas and liver pathology
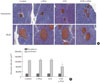
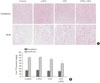
Histopathological analysis of the pancreas found db/db mice to have significantly increased islet size when compared to control nondiabetic heterozygous mice (Fig. 5A and B); however, islet size in the GTE+γ-PGA diet db/db mice was maintained at a relatively smaller size compared with the control diet db/db mice. Additionally, db/db mice had significantly increased liver size (relative to BW) compared to control nondiabetic heterozygous mice (data not shown). Histopathological analyses showed that these mice also had a significantly higher incidence of fatty liver, whereas GTE+γ-PGA diet db/db mice had reduced incidence of fatty liver compared to control diet db/db mice (Fig. 6A). Triglyceride content was significantly decreased in GTE+γ-PGA diet db/db mice compared to control diet db/db mice (Fig. 6B).
DISCUSSION
The NMR results of this study have shown that γ-PGA can interact with EGCG, a major GC of GTE. This possible complex formation in the intestine may delay the absorption of GCs of GTE that ameliorate BW gain and glucose intolerance in db/db mice in the 4-week drug treatment. That the regimen was not effective in nondiabetic mice may indicate that it does not have significant side effects. GTE+γ-PGA treatment did not change food intake and hypothalamic NPY expression, suggesting that the drug does not act on the hypothalamic appetite center; rather, it reduced the uptake of calories from the intestine. In contrast to other therapeutic drugs inhibiting gastrointestinal absorption of glucose, such as α-glucosidase inhibitors, a novel advantage of GTE for reducing gastrointestinal calorie intake may be based on its inhibition of both glucose and cholesterol absorption. Therefore, GTE+γ-PGA regimen may be a good candidate for supportive prevention and treatment of both obesity and T2DM with relatively few side effects.
This study has shown that the administration of GTE alone did not significantly ameliorate BW gain and glucose intolerance in a 4-week treatment at the tested concentrations. We have previously confirmed that GTE, when simultaneously administered with glucose, reduces postprandial glucose elevation [7], which is consistent with many other reports [8,16-18]. However, as shown in this study, the effect of GTE can be reversed when GTE is administered approximately 30 to 60 minutes before glucose loading, implying its ambivalent effects on BGLs depending upon the time between the administration of GTE and glucose [7]. In the present study, GTE was always supplied mixed with chow diet. Adverse effect of GTE on glucose intolerance may appear if GTE is given to animals between meals. In contrast to this study, EGCG given at higher concentrations could reduce food intake and BW even in normal rats by reducing appetite through the hypothalamus [19]. Various complications, however, were detected at such high concentrations of EGCG, one of which is antiandrogenic effect of EGCG [20].
Intake of Korean chungkookjang and Japanese natto has long been known to be effective in reducing blood cholesterol levels and increasing insulin sensitivity and cellular antioxidant activity [21-25]. The main active components of chungkookjang and natto are thought to be isoflavones [26]. γ-PGA, another component of chungkookjang and natto, was also reported to have antioxidant activity [27]. Although some reports have discussed the capacity of high molecular weight γ-PGA (>2,000 KDa) to reduce BW and blood cholesterol levels in animals and humans [26,28], the present results do not provide antiobese and antidiabetic effects of γ-PGA alone (MW: 2,000 KDa) in db/db mice. The reason may be the difference in not only the molecular weight, but also the treatment duration with γ-PGA. The short duration of experiment of this study may not detect the full effect of γ-PGA. The effect of prolonged treatment with γ-PGA alone remains to be further determined.
The beneficial effect of GTE+γ-PGA on glucose tolerance appears to result from lower BW gain and decreased body fat mass, a similar mechanism to α-glucosidase inhibitors [29]. Like α-glucosidase inhibitors, GTE+γ-PGA may cause gastrointestinal side effects, such as diarrhea and soft feces. However, GTE+γ-PGA can prevent the absorption of simple sugars by inhibiting glucose transport, while α-glucosidase is unable to do so. An additional benefit of GTE+γ-PGA may be the inhibition of cholesterol absorption from the alimentary tract [30]. GCs appear to be responsible for the inhibitory effect, primarily through the inhibition of mixed micelle formation in the intestinal lumen [30].
In conclusion, this study has shown that GTE+γ-PGA treatment can significantly alleviate BW gain and the development of glucose intolerance in db/db mice, probably through the action of intestinal GTE. This combined treatment is much more effective than treatment with either GTE or γ-PGA alone. Therefore, a GTE+γ-PGA regimen may be a promising preventative and therapeutic candidate for obesity and obesity-related metabolic disorders, such as T2DM.




 XML Download
XML Download