

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Metformin is the preferred insulin-sensitizing drug in obese diabetes patients due to it suppresses appetite and reduces body weight [1-3]. Metformin acts in the liver and muscle by activating AMP-activated protein kinase (AMPK), and is known to increase the activity of insulin [4-6]. Metformin reportedly activates AMPK through a series of sequential steps through phosphoinositide-3-kinase (PI3K), protein kinase C (PKC)-ζ, and liver kinase B1 [7].
Appetite suppression by metformin is more prominent in obese model with hyperleptinemia than in lean control [8], and therefore has more clinical significance. However, the mechanism has not yet been completely investigated. Although several reports state that metformin suppresses appetite by increasing the insulin sensitivity of peripheral tissues [2,3], other studies suggest that additional factors affecting the appetite are active in the brain. Kim et al. [8] reported an increase in the phosphorylated signal transducer and activator of transcription 3 (pSTAT3) in the hypothalamus after metformin injections. Billa et al. [9] reported a change in luteinizing hormone and prolactin as a result of metformin action in the pituitary gland of patients with polycystic ovary syndrome. Vigersky et al. [10] reported biochemical or clinical thyroid-stimulating hormone suppression without thyroid hyperactivity in patients who were taking metformin, and discussed the possibility of metformin affecting the brain directly. However, effects on appetite suppression through direct in vivo metformin action in the brain have not yet been reported.
Currently, metformin is known to increase the only identified target substance, AMPK, in muscles and the liver. However, the mechanism of action in the hypothalamus has yet to be identified [11]. Recently, Chau-Van et al. [12] reported that metformin caused a decrease in pAMPK concentration in primary cultures of hypothalamic cells. Ropelle et al. [13] reported that orally administered metformin increased pAMPK in the hypothalamus. Overall, results for the mechanism of metformin in the hypothalamus are conflicting, thus clarification is necessary.
The present study was performed in order to investigate the direct effects of metformin on the brain, particularly in the hypothalamus, and the mechanisms of those effects by delivering metformin through a cerebroventricular injection in order for changes in food intake to be observed. The appetite-regulating proteins of the hypothalamus were also analyzed.
METHODS
Experimental animals
Male Sprague-Dawley rats weighing ~250 g were purchased from Samtako (Seoul, Korea). All experimental animals were kept by animal specialists from the Yeungnam University College of Medicine. Animals were given roughly 1 week to acclimate before they were used in the experiment. The rats were exposed to 12-hour light/dark cycles. Light exposure began at 7:00 AM and ended at 7:00 PM. The animals were cared for in accordance with the principles of the Guide to the Care and Use of Experimental Animals of the Yeungnam Medical Center.
Experimental design
Experiment 1
To investigate whether metformin acts directly on the brain to suppress appetite, various concentrations of metformin (0, 3, 30, or 300 µg) (Sigma Chemical Co., St. Louis, MO, USA) were injected into the left cerebral ventricle and the food intake of the rats was measured and evaluated for 24 hours.
The dosages of metformin were determined by a modification of previous report [14] that metformin (320 mg/kg/day) caused an increase in in vivo skeletal muscle glucose consumption.
Experiment 2
To determine the mechanism of metformin, varying concentrations were injected into the cerebral ventricle (0, 3, 30, or 300 µg). After 1 hour, the hypothalamus was extracted, the amount of the diet control signals pAMPK, pSTAT3, and PI3K and the expression of proopiomelanocortin (POMC) and neuropeptide Y (NPY) mRNA were measured and compared.
Placement of a chronic intracerebroventricular (ICV) catheter
The rats were anethesized with intra-peritoneal injections of 25 mg/kg of tiletamine and zolazepam (Zoletil®; Virbac, Carros, France) and 8 mg/kg of xylazine hydrochloride. After anesthesia, rats were fixed with stereotaxic instruments, an incision was made in the flesh along the midline, and a hole was made 1.3 mm posterior, 1.9 mm lateral, and 3 mm deep from the bregma. A cannula made by 21-gauge syringe needle was inserted into the lateral ventricle and the orifice was occupied with a wire sized to fit inside the cannula. The brain cannula was secured to the surface of the skull using a jewler's screw and acrylic dental cement. Drug injections were given 1 week after surgery to allow for recovery.
ICV administration of drug
In order to observe short-term dietary intake, the wire placed earlier was removed in freely moving rats, a pre-made syringe needle that matched the length of the cannula was inserted, and an injection was given over a period of 3 minutes. The syringe was removed 3 minutes after the injection. The injection consisted of 5 µL of saline solution with 0, 3, 30, or 300 µg of metformin. Injections were given at 4:30 PM in order to measure food intake activity at night. At 3, 7, 16, and 24 hours after the injection, food intake was measured.
In order to observe signal transduction pathways in the hypothalamus after metformin was administered, rats underwent lateral cerebroventricular intubation while anesthetized after which the aforementioned procedure with different amounts of metformin (0, 3, 30, or 300 µg) was administered. Metformin solution was matched with 5 µL applied over 3 minutes and removed 3 minutes afterwards. After 1 additional hour, the hypothalamus was removed and examined.
Sample collection
At the end of the experiment, the rats were injected with a double dosage of anesthetic prior to 9:00 AM. The hypothalamus was then harvested, stored in liquid nitrogen, and analyzed using the method employed by Kim et al. [15]. Briefly, the hypothalamus, which is located at the base of the brain along the optic nerve margin and typically 3 mm deep, was collected. The extracted hypothalamus was bisected down the centerline, and was used for analysis of proteins and gene expression.
Western blot
Appetite-related neural peptides in the hypothalamus were analyzed using a Western blot. STAT3, pSTAT3, AMPK, pAMPK, and PI3K were targeted for the analysis of signal transduction protein products. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal standard. After 30 µg of hypothalamus protein was separated on a 10% polyacrylamide gel, the nitrocellulose membrane was removed, antibodies for each protein (Cell Signaling, Danvers, MA, USA) were fixed, the membranes were washed, and secondary antibodies prepared. After a combined blot was presented using an ECL kit (Amersham Biosciences, Little Chalfont, UK), quantified analysis was performed using Scion Image Software (NIH, Bethesda, MD, USA). Each blot removed attached antibodies, and another antibody fixing method was performed.
Real-time polymerase chain reaction (PCR)
Gene expression of the hypothalamus was analyzed using real-time PCR. To summarize the process of RNA extraction, 1 mL TRI-reagent (Sigma Chemical Co.) was added to the hypothalamic tissue and homogenized using ultrasound. Next, 200 µL of chloroform was added and the mixture was shaken vigorously. The sample was left at room temperature for 5 minutes to allow for the reactions to occur. Afterwards, the sample was centrifuged at 13,200 rpm for 15 minutes at 4℃. An equal volume of isopropanol was added to the separated supernatant and left at room temperature for 10 minutes. The sample was then centrifuged at 13,200 rpm for 10 minutes at 4℃ to pellet precipitated RNA. The precipitated RNA was washed with 1 mL of cold 75% ethanol, centrifuged at 13,200 rpm at 4℃ for 5 minutes, and dried at room temperature. Finally, the remaining RNA was dissolved in 30 µL RNase free water and stored at -70℃. RNA concentration was measured using a Nano-drop (Thermo, Wilmington, MA, USA) between 260 and 280 nm wavelengths. Briefly, to amplify the RNA the isolated 1 µg RNA was reacted for 5 minutes at 70℃, oligo (dT) primer was added to Prime RT Premix (iNtRON, Seongnam, Korea) and was kept at 42℃ for 60 minutes and 70℃ for 10 minutes to produce the cDNA. Afterwards, FastStart Taq DNA polymerase (LightCycler FastStart DNA Master SYBR Green I; Roche, Mannheim, Germany), 3 mM MgCl2, and 0.4 µM of primer was added and amplified in a LightCycler 1.5 Instrument at 95℃ for 10 minutes. After the process was completed, the sample was denatured at 95℃ for 10 seconds, annealed at 54℃ to 60℃ for 5 seconds, and extended at 72℃ for 4 to 9 seconds. This process was repeated 45 times. Once the reactions were completed, to confirm the specificity of the amplification, the temperature was decreased to 65℃, and gradually increased by 0.2℃/sec until the temperature reached 95℃. The PCR products were then analyzed. β-actin was used as an internal standard. The primer sequences used are shown in Table 1.
Statistical analysis
Values were expressed as mean±standard error. For the cumulative dietary intake relative to metformin dosage concentration, the concentration dependence was evaluated as the P trend qualification. The comparison of the amount of expression of signaling proteins and neuropeptides in each group was performed using a one-way ANOVA test, and a post-hoc test was performed using Duncan.
RESULTS
In order to observe how metformin reduces appetite by acting directly on the brain, a long-term catheter was installed which administered metformin. The 24-hour cumulative food intake was recorded and is shown in Fig. 1. The cumulative food intake appeared to decrease as metformin concentration increased, reflecting concentration dependence.
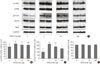
In order to examine the appetite suppressing mechanism of metformin, the hypothalamus was collected 1 hour after an injection was made into the cerebral ventricle and signaling molecules involved with appetite regulation were analyzed. pAMPK increased in the hypothalamus when 3 µg of metformin was applied, but there was no further change as metformin dosage increased. Phosphorylation of STAT3 increased as a result of metformin, but further changes did not occur as a result of increasing metformin dosage. PI3K did not show any specific changes (Fig. 2).
The effects of metformin on the appetite controlling hypothalamic peptides POMC and NPY were observed. POMC mRNA, which is known to suppress appetite, increased in the presence of metformin but was not dependent on concentration, and NPY mRNA did not decrease in the presence of metformin (Fig. 3).
DISCUSSION
It has been suggested that the appetite suppressing mechanism of metformin can be achieved by being applied directly to the brain [8-10,12]. However, no studies have been published investigating the effects on appetite suppression of metformin administered in vivo directly to the brain.
In the present study, in order to observe the effects of metformin on the brain, specifically the hypothalamus, metformin was injected directly into the cerebral ventricle. The decrease in food intake was examined for 24 hours after metformin was administered in a dose-dependent manner (Fig. 1). In order to determine whether the loss of appetite due to metformin was reversible, food intake was analyzed 48, 72, and 96 hours after metformin injections (data not shown). Food intake returned to normal after 24 to 48 hours after 3 and 30 µg of metformin was administered, and after 72 to 96 hours after 300 µg of metformin was administered. These results imply that metformin can act directly to suppress appetite in the brain.
In order to clarify the central appetite-suppressing mechanism of metformin, metformin was injected into the cerebral ventricle, the hypothalamus removed, and appetite-related signaling molecules (AMPK, STAT3, PI3K) levels and neuropeptide (POMC and NPY) expression were analyzed.
AMPK is a metabolic sensor which detects cellular energy and reacts appropriately. When AMP increases, activation by phosphorylation occurs [16,17]. The increase in pAMPK in skeletal muscle increases energy production during metabolism. However, an increase of pAMPK in the hypothalamus increases appetite. Conversely, the reduction of pAMPK is known to suppress appetite [18,19]. The appetite-increasing effect of phosphorylated AMPK is known to occur through increased NPY, which is blocked by treatment of compound C, a known AMPK inhibitor [12]. AMPK is also controlled by leptin; however, leptin activates AMPK in skeletal muscle and is known to reduce pAMPK in the hypothalamus [11]. In the present study, metformin injections appeared to increase pAMPK. These results were consistent with the results reported by Ropelle et al. [13] that metformin injections caused an increase in pAMPK in cancer cell inoculated rats. However, there was a difference with the in vitro results reported by Chau-Van et al. [12]. The differences were not only due to the difference in the in vitro and in vivo environments, but Chau-Van et al. [12] examined the metformin's effect on a glucose-deprived in vitro condition which elevates pAMPK markedly. In the aforementioned study, when metformin was administered to hypothalamic neurons when glucose levels were normal the pAMPK inhibition effect was not significantly apparent [12]. ICV metformin treatment suppressed appetite, but failed to reduce pAMPK in this study, suggest that the AMPK pathway has little association with reduced appetite or metformin under normal conditions. Further research on the effects of metformin on the hypothalamus in patients with hyperglycemia or hyperlipidemia is required.
STAT3 is activated by phosphorylation through tyrosine kinase activity associated with leptin receptor autophosphorylation [20]. The increase in phosphorylated STAT3 leads to the increase of POMC and allows for appetite suppression [21]. In the present study, pSTAT3 increased starting at 3 µg of metformin, but when the metformin dosage was increased to 300 µg, pSTAT3 did not increase any further. Although the appetite-suppressing effect of metformin is related with pSTAT3 elevation at least in part, since there was not shown any concentration-dependent increase of pSTAT3 by metformin, this was not considered to be the primary action mechanism. PI3K, another pathway that carry appetite and related signals, activates POMC neurons and is also known to contribute to insulin action [22,23]. In the present study, hypothalamic PI3K did not show any particular change after metformin administration, suggesting detailed studies on mechanisms are required due to the possibility of several forms of action of metformin on the brain. If a new appetite control pathway can be discovered, it could possibly be manipulated for treatment of obesity.
In the present study, upon metformin injection, through changes in neuropeptides, POMC mRNA increased, but NPY mRNA did not appear to decrease. Changes in POMC mRNA had a tendency to be very similar to changes in pSTAT3. Changes in NPY mRNA had a tendency to be similar to changes in pAMPK observed in the present study and were consistent with reports from other studies [12,24]. Rouru et al. [25] suppressed appetites in Zucker rats using metformin, but reported that there was no reduction in NPY. Taken together, the appetite suppression action of metformin may not be a result of the inhibition of pAMPK and NPY.
In conclusion, the result of injecting metformin into the lateral ventricle of rats produced concentration-dependent appetite suppression, and the direct suppression of appetite through metformin to the brain could be confirmed. One hour after metformin was injected into the lateral ventricle, hypothalamic pSTAT3 increased, but concentration dependence was not observed. However, metformin did not decrease the amount of hypothalamic pAMPK. Thus, inhibition of appetite by metformin is thought to be partially due to the increase in pSTAT3, although other studies investigating mechanisms besides STAT3, AMPK, and PI3K are considered necessary.

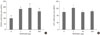

 XML Download
XML Download