

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Several in vitro models focusing on cells or tissues have been described and used to induce traumatic brain injuries (TBI). These models include the effects of transaction,13,29,37) compression,4) acceleration,24) hydrostatic pressure,38) hydrodynamic forces21) and stretch.11) In comparison with cultured cells, cultured or sliced organotypic brain tissues contain heterogeneous populations of cells and have the advantage of maintaining complex three-dimensional connections. Acute hippocampal slices (AHSs) have a long history of successful application in neural science investigations, and the organotypic slice culture (OTC), cultured on semiporous membranes, has become popular for studying mechanisms of brain damage. Because these fragments of nerve tissue are very small, it is not easy to induce reproducible, targeted damage. Previously described damage methods have, therefore, been limited to weight drop,1,4,33) barotraumas38) and substrate strain.27,28)
The centrifuge is commonly used in laboratories to apply a known amount of centrifugal force to target tissues. Tissues under acceleration undergo an increase in pressure, and this generates strain and transformation. Thus, centrifugal forces applied to tissues provide a reliable in vitro model of static compression injury, producing reproducible damage without special equipment. We have investigated the benefits of this model.
Materials and Methods
Acute hippocampal slice preparation
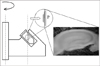
Male Sprague Dawley rats, 4-6 weeks of age and weighing 100-150 g, were anesthetized with 3% halothane and decapitated. The brains were rapidly removed and placed in artificial cerebrospinal fluid (aCSF), chilled to 0-4℃ and oxygenated with a mixture of 95% O2 and 5% CO2. The hippocampi were removed from each rat brain and cut into 450 µm thick transverse slices using a McIlwain tissue chopper. Slices were incubated in vials of continually oxygenated aCSF at 30±1℃ before use. The aCSF consisted of 124 mM NaCl, 3.0 mM KCl, 2.0 mM CaCl2, 2.0 mM MgCl2, 1.2 mM NaH2PO4, 26 mM NaHCO3 and 10 mM d-glucose. Aeration with 5% CO2 in O2 resulted in an aCSF pH of 7.4 ± 0.05. After a 90 min recovery period, each slice was laid on a piece of lens cleansing paper (Cat. No. 2105 862, Whatman) for later handling using forceps (Figure 1). A blind protocol was used to assign slices to either a sham-injured or injury group, and for analyzing the results. All animal procedures were reviewed and approved by the Korea University Institutional Animal Care and Use Committee, and complied with guidelines outlined in the Korean Academy of Medical Science's Guide for the Care and Use of Laboratory Animals.
Organotypic hippocampal slice culture
Seven-day-old Sprague Dawley rat pups were anesthetized with halothane and plunged into a 70% alcohol solution. They were decapitated, and transverse hippocampal slices (400 µm) were taken and kept in an ice-cold stabilization medium for approximately 30 min. The medium contained 50% minimal essential medium (MEM) with no bicarbonate or glutamate, 50% calcium- and magnesium-free Hanks' balanced salt solution (HBSS), 7.5 mM glucose and 20 mM N-2-hydroxyethyl piperazine-N'-2-ethanesulfonic acid (HEPES), at pH 7.15. Following stabilization, slices were separated and transferred to 25 mm inserts, each containing a porous (0.4 µm) Millicel-CM sterile tissue culture membrane (Millipore, Bedford, MA, USA). Inserts, each with six equally spaced hippocampal tissue slices, were transferred to six-well culture trays with 1 mL of growth medium in each well. The medium consisted of 50% MEM, 25% horse serum, 25% Earl's balanced salt solution with glucose and HEPES, 5,000 units/mL penicillin G and 50 mg/mL streptomycin sulfate, adjusted to pH 7.3. Slices were cultured for 14 days at 36.5℃ and 100% humidity, in an atmosphere of 95% air and 5% CO2 and fed twice weekly by 50% medium exchange. To minimize culture variability, each set of control and experimental OTCs within a study were cultured on the same day. The inserts were blindly assigned to either a control, sham-injured or injury group; each plate also contained a control insert representing the status of the plate under normal culture conditions during the period of the experiment.
Centrifugal injury application
We used two different methods to apply centrifugal stress. OTCs on their 25 mm inserts were simply placed at the bottom of a sterile centrifuge bucket in a swinging rotor. This type of centrifuge has a slightly longer period of acceleration, with experimental forces lasting up to 3 min (including the acceleration and plateau phases). For AHSs we used an angled rotor with a rapid acceleration phase. To arrange AHSs perpendicular to the axis of rotation in the angled rotor, melted paraffin was poured into an Eppendorff tube and the flat paraffin surface so formed was positioned parallel to the rotational axis (Figure 1). Sham-injured slices were subjected to all procedures except centrifugation.
Morphometric study of the CA region
Fluorescence microscopy was used to analyze results from some AHS experiments. First, the nuclei in the pyramidal layer at the slice surface killed during slice preparation were labeled by cultivating the slices in aCSF containing 10 µg/mL propidium iodide (PI) for 30 min prior to injury. The slices were photographed at 40× magnification before injury and again under the same conditions after centrifugal acceleration. The area of the cornus ammoni (CA) field between the hilus and CA1 was measured using image analyzing software before and after injury, and the extent of any change expressed as a percentage.
CA1 population spike recordings
After a 30 min recovery period, the AHSs backed by the lens paper were transferred to a submerged 0.12 mL recording chamber. The superfusing aCSF (2 mL/min) was continually aerated and its temperature was kept at 32-33℃. Evoked extracellular population spikes (PSs) were recorded in the stratum pyramidale of CA1 following electrical stimulation of the Schaffer collateral commissural pathway. The spikes were recorded with glass micropipettes filled with 150 mM NaCl (2-5 MΩ). The field potentials were amplified, filtered and recorded on-line with a computer interface. Stimuli (50 µs in duration) were delivered at 0.05 Hz through bipolar platinum-iridium electrodes with tip diameters of 25 µm, placed in the stratum radiatum. The minimum strength necessary to elicit a maximal orthodromic population spike (PS) was applied and the PS observed for at least 30 min.
Measurement of cell death
Viability and cell death in OTCs was examined by the exclusion of fluorescent dye using an inverted fluorescent microscope after culturing the slices in a medium containing 7.5 µM PI. Cultures displaying a high initial PI uptake were excluded from further experiments. The cell death index derived from the fluorescent intensity measure was normalized to the values obtained after the slices were exposed to an ambient temperature of 4℃, which results in maximal cell death. Pixel intensity within a selected area was averaged to enable statistical comparisons to be made. Some OTCs were exposed to a secondary injury between 12 and 24 hr after the initial injury. Hypoxic injury was applied by placing OTCs in a 37℃ incubator gassed with 95% N2/5% CO2. Exposing OTCs to 10 mM glutamate, or depriving them of serum, induced excitotoxic and apoptotic cell death. A second impact was delivered by repeating the centrifugation at an identical strength. All slices from the same plate were discarded from the study if OTCs in the control insert showed more than slight PI (-5%) fluorescence at the end of the experiment. All recorded data were expressed as mean±standard errors. Two sample t-tests were carried out for the comparisons between the groups. The results with p values less than 0.05 were considered to be significantly different.
Results
Morphometric observations
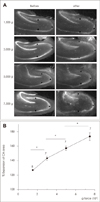
The morphological changes in acute hippocampal slices after centrifugal acceleration are characterized by flattening of slices, expansions of subfields, splits in natural fissures, cleavages of cellular layers, and loss of tissue (Figure 2A). The transformation was particularly severe within the dentate gyrus (DG), with the disruption of tissue making morphometric assessment of the degree of expansion unreliable. The field expansions were distinctive even in the hilar region. In CA regions the slices retained their original morphology well, with centrifugal forces applied to an AHS for 60 sec showing a graded effect on tissue expansion (Figure 2B).
Field potential recordings
The PS amplitude measured 30 min following sham injury (at 1 g) on the control slice was 6.6+/-0.5 mV. Increasing the force for 60 s gradually reduced the amplitudes of PS with the minimum level (<10%) being recorded at 5,000 g (Figure 3A). We arbitrarily divided the force applied for sham injury to minimum level into five levels to obtain a graded injury (500, 1,500, 2,300, 3,000, and 5,000 g). The mean PS amplitudes measured at 30 min after injury were significantly different between these levels. At 2,300 g, a 20 s step increase in duration also revealed statistical differences in PS values (Figure 3B).
Cell Death in OTCs
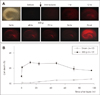
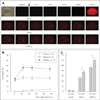
Four days of posttraumatic observations revealed that the peak PI uptake developed within the first 24 hr after trauma, and slightly decreased thereafter (Figure 4). During the observation period, cell death in sham injured cultures was minimal. The most unexpected observation was a widespread increase in PI uptake of about 25% seen at 1 hr in the OTCs subjected to either slight (300 g) or moderate (3,000 g) centrifugation. Twelve hours after injury, diffuse PI uptake was less well defined and delayed degeneration began in the neuronal layers, usually in the inner blade of the dentate gyrus (Figure 5). Three different strengths of centrifugal force (300 g, 1,000 g, and 3,000 g) resulted in graded peak uptakes of PI. Cell death was prominent in both the dentate gyrus (DG) and CA1 in more severely damaged OTCs, but the progression of cell necrosis was minimal up to the end of the experiment, regardless of the force intensity. With forces stronger than 5,000 g, OTCs frequently became detached from the porous membrane, making measurements of identical areas impossible. Secondary exposure to excitotoxicity (glutamate, 10 mM for 30 min, n=12), or hypoxia (for 60 min, n=12) led to increased cell death in the neuronal layers of both sham-injured and injured OTCs, whereas serum deprivation (for 24 hr, n=12) or additional impact (300 g, n=12) was more likely to produce diffuse patterns of cell death in traumatized OTCs (Figure 6).
Discussion
Static compression of nervous tissue has been a classical model of traumatic injury for many years. One of its limitations is that the mechanism of dynamic impact is not included. Nevertheless, by adjusting both the extent and duration of compression, controlled, graded injuries can be produced. In vivo spinal cord models include clip compression30,31) and subdural balloon inflation,25) with acute subdural compression7,20) used for the intact brain. Clip compression was used in vitro on a strip of dorsal column from the spinal cord.14) The cylindrical shape of optic nerves and spinal cord strips facilitates the application of calibrated compressions. In contrast, thin, wide brain slices require more delicate and sophisticated techniques because they lack supportive structures. Limited access time without perfusion is another potential problem with in vitro experiments: only a few minutes spent adjusting the apparatus and sample tissue can destroy the tissue. The technique described here, however, does not suffer from these limitations, and in addition provides easy, reproducible adjustment of the applied force. Special training is not required, which minimizes variations in the injury applied. The major advantage of this injury system is its versatility. A single apparatus suitable for producing injury in both acute preparations and OTCs has not been described previously. Although we have only tested it on hippocampal slices, it should work well with other tissue preparations. The force applied to the tissue with this technique is simply expressed in gravitation units, making direct comparisons with data from other experiments possible.
The effect of acceleration on the brain depends on its duration and strength, and on the brain's biophysical properties. Studies performed in the 1950s that exposed human volunteers to up to 40 g of sagittal deceleration in a linear decelerator did not produce lasting neurological complaints or evidence of permanent brain damage.36) However, blunt trauma sustained in falls from standing positions can generate 100 g of force, and produce fatal head injuries.16) The brain generally is better able to tolerate slow increases in acceleration, even though the total amount of strain is identical. Wearing a helmet increases the duration of impact and lowers the peak force, as shown by accelerometer measurements.22) Helmeted racing car drivers have had no brain injuries from accidents involving head decelerations of over 200 g, and jet aircraft test pilots have been subjected to up to 450 g without concussion.34) Small tissue slices offer several advantages over whole human brains in the study of mechanical injury. The ratio of brain weight to surface area is low in small brain and tissue samples, and therefore a given impact deceleration will be spread over a correspondingly greater area. The acceleration required to produce injury in brains with similar properties and shapes should be inversely proportional to the two-thirds power of their mass. The acceleration required to produce an injury similar in extent in whole human would, therefore, need to be 400-500 times greater for 450 µm slices. The functional loss in the human brain assumed to be caused by a dynamic impact of 60 g would require up to 30,000 g in brain slices. Small size is one reason why woodpecker brains, weighing 1.25-3.95 g, can withstand repeated impact forces of the order of 1,000 g during impact drilling.26) The single impact acceleration threshold to produce concussion in dogs is between 250 g and 500 g.18) Even with same acceleration applied in a well-designed centrifugal system, tissue transformation is affected by other physical conditions. One variable is the density and elasticity of the slices, especially in AHS preparations, which are affected by temperature and incubation time during preparation and storage. Tissue slices gradually become swollen due to surface autolysis and water uptake increasing the damage; this begins within minutes of preparation.5) Testing the slices at the same temperatures and incubation duration can minimize this damage. Even with identical tissue densities and elasticities, the pressure effects vary within the thickness of the slice. Generally, structures in the middle are not deformed by ambient gravity. However, the increased pressure from overlying tissue is multiplied by the g force such that, in thicker slices, the deeper regions are more deformed by centrifugation. Therefore, measurement of tissue at the same depth from the top surface is a key to obtaining reproducible results. For this reason, the AHS data (usually recorded 150 µm from the surface) are more consistent than images from OTCs obtained from the bottom of the slices. Additional injury caused by the stretching of the suspended porous membrane in the inserts during centrifugation might also be responsible for the observed variation. This membrane is elastic and is attached at the wall, just above the bottom of the insert. It is at first stretched downwards during centrifugation until it comes into contact with the floor of the apparatus. Such initial, limited strain and shear between the slices and the stretched membrane may be responsible for the widespread uniform intensity in PI uptake measured at 1 hr, regardless of the force applied.
The prominent injuries observed in this study, especially in the medial blade of the dentate gyrus in both AHSs and OTCs, are striking. Although all areas of the slice are equally exposed to the force in this experimental technique, the circular configuration of the cell layers in the dentate gyrus may offer a localized area of relative weakness to shear forces. In vivo studies have reported obvious differences in the pattern of cell death in various models.15) Although studies using cortical impact or fluid percussion injury have demonstrated significant neuronal cell loss in the CA2/CA3 and hilar regions of the dentate gyrus3,17,35) other investigators have noted dentate hilar cell loss with no significant loss in CA1, CA2, or CA3 using the same paradigm.6,23) Neuronal loss in the dentate gyrus is confined to the hilus and delayed in onset, but it is probable that dentate granule cells showing early32) and delayed23) posttraumatic hyperexcitability are also susceptible to trauma. Similarly, the pyramidal cells in CA1 with posthypoxic hyperexcitability10) have been shown to be selectively vulnerable to hypoxia.2,19) Although there is no evidence, perhaps the early, five-fold increase in the number of progenitor cells in the dentate gyrus12) plays a role in replacing neurons lost in CA3, and possibly in the dentate gyrus itself. We also note the difference in the temporal pattern of posttraumatic cell death from previous models1,9) in which cell death is greatest at 3-4 days. We have no explanation for these differences, but our findings agree with previous in vivo studies in which neuronal loss reached a maximum in the first day post trauma, and remained near that level subsequently.3,8) The known selective vulnerability to hypoxia in CA1 and excitotoxicity in CA1 and CA3 was reproduced in our trauma model. However, secondary impact or apoptosis induced by serum deprivation produced a more widespread cell death in our study, suggesting that these insults might have a tendency to kill glia as well as neurons in the posttraumatic state.
In conclusion, we showed novel compressive model of neural injury. It may be useful for neurotrauma research because it is accessible, easily adjustable, and reliable.


 XML Download
XML Download