

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Saliva is a mixed glandular secretion that coats and protects the oral cavity and is vital to the maintenance of healthy teeth and soft tissues, such as the gingiva. Salivary fluid formation is primarily driven by Ca2+-activated, apical efflux of chloride into the lumen of the salivary acinus [1]. Native chloride channels are normally quiescent, but become highly activated when intracellular Ca2+ levels rise above 100 nM, as occurs following muscarinic stimulation. In order to maintain electroneutrality, Na+ follows the Ca2+-driven Cl- and together with the osmotically-obliged water, they form the isotonic luminal fluid in the salivary acini before being transported and modified by the salivary ducts and ultimately, excreted into the oral cavity [1]. When expressed heterologously in mammalian cells, a recently discovered protein, anoctamin1 (TMEM16A), recapitulates the electrophysiological properties of the endogenous calcium-activated chloride currents in the salivary acinar cells, which are activated by submicromolar levels of intracellular calcium, are outwardly rectifying, and are blocked by inhibitors such as niflumic acid, and 4,4'-diisothiocyanato-stilbene-2,2'-disulfonic acid (DIDS) [2,3,4].
Anoctamin1 is the founding member of a family of ten transmembrane proteins with similar predicted architecture, comprising of eight transmembrane segments with cytosolic N- and C-termini [2,3,4]. In addition to its role in exocrine secretion, the chloride channel function of anoctamin1 plays an important physiological role in gut motility, airway function, smooth muscle contraction, and nociception [5]. With the exception of the anoctamin2 and anoctamin6 proteins, the other anoctamin isoforms do not form functional ion channels in plasma membranes. Despite their unclear roles in cell function, the evidence for disease-associations are compelling for some anoctamin proteins: mutations in anoctamin5 leads to several musculoskeletal disorders including gnathodiphyseal dysplasia and proximal limb-girdle muscular dystrophy [6,7]; mutations in anoctamin6 are implicated in defective platelet function and blood clotting in Scott syndrome [8]; and mutations in anoctamin10 have been linked to spinocerebellar ataxia [9]. The diverse spectrum of human diseases stemming from anoctamin mutations suggests an unusually wide array of physiological functions, in addition to the role of these proteins in mediating Ca2+-activated Cl- conductance in cells.
In order to better understand the role of anoctamin proteins in salivary gland function, it is essential to identify which of the anoctamin isoforms is present in the tissue. Here, we report that anoctamin1 is the main isoform found in the mouse submandibular gland, but multiple anoctamin isoforms and splice variants thereof are also expressed, including anoctamin5 and anoctamin6.
MATERIALS AND METHODS
RNA isolation and reverse-transcription polymerase chain reaction
Total RNA was extracted from four eight-weeks-old female mouse (B6D2F1) submandibular glands after polytron homogenization of the tissue using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. First-strand complementary DNA (cDNA) was prepared from 2 µg of RNA isolated from two animals using 1 pmol of oligo (dT)20 primer and 200 units of M-MLV reverse transcriptase (Promega, Madison, WI, USA) for each 25 µL reverse-transcription reaction. The first-strand cDNA synthesis was repeated using RNA extracted from two different animals. The expression of all anoctamin isoforms (anoctamin1-anoctamin10) was determined by reverse transcription-PCR (RT-PCR) using specific oligonucleotide primers (Table 1). i-Taq DNA polymerase (Intron, Seongnam, Korea) and Phusion High-Fidelity DNA polymerase (Finnzymes, Keilaranta, Finland) were used to amplify PCR products in the screening and the full-length cloning, respectively using routine molecular biological methods. A touch-down PCR protocol was performed on C1000 Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA) as follows: 95℃ five minutes, 31 cycles at 95℃ for 30 seconds, 70℃-55℃ for 30 seconds (reduction of 0.5℃ per cycle), 72℃ for three minutes, 10 cycles at 95℃ for 30 seconds, 55℃ for 30 seconds, 72℃ for three minutes, followed by a final step at 72℃ for five minutes.
Full-length anoctamin cloning
For full-length cDNA cloning, the PCR amplification products were size-separated on 0.8% agarose gel, cut out, and purified using Dual PCR Purification Kit (Bionics, Seoul, Korea) and subcloned into the pCDNA3.1(+) mammalian expression plasmid, and sequenced to confirm the fidelity of the PCR-amplified sequence against the data from Genbank (Anoctamin1-NM_178642.5; Anoctamin2-NM_153589.2; Anoctamin3-NM_177694.5; Anoctamin4-NM_178773.4; Anoctamin5-NM_177694.5; Anoctamin6-NM_175344.4; Anoctamin7-NM_207031.1; Anoctamin8-NM_001164679.1; Anoctamin9_178381.3; and Anoctamin10-NM_133979.2).
RESULTS
Anoctamins in the submandibular gland
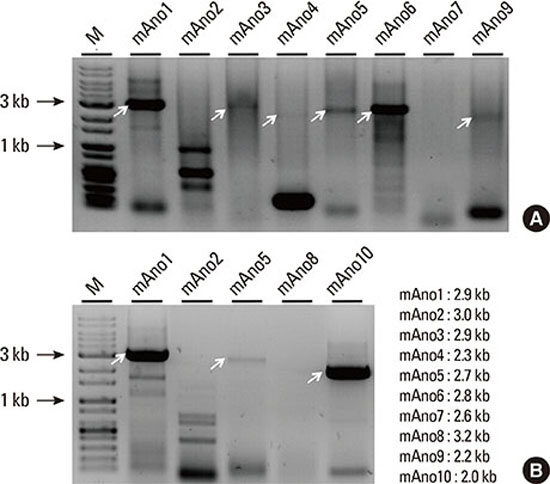
In order to determine which of the anoctamin isoforms were expressed natively in the mouse submandibular gland, RT-PCR was performed from total RNA. A pair of oligonucleotide primers specific for each anoctamin isoform was designed to amplify the entire open-reading frame, based on the mouse nucleotide sequence found in the Genbank database. Each primer was engineered with a unique restriction enzyme to facilitate directional subcloning (Table 1). Complementary DNA sequences corresponding to full-length anoctamin1, anoctamin10, anoctamin6, anoctamin5, anoctamin3, anoctamin9, and anoctamin4 (rank order of transcript abundance) were amplified, and purified for subcloning (Fig. 1). Although the results were not strictly quantitative, the anoctamin1 isoform appeared to be the most abundant followed by anoctamin10 and anoctamin6. Despite multiple RT-PCR trials, full-length anoctamin2, anoctamin7, and anoctamin8 could not be amplified, suggesting that these isoforms are not expressed in the mouse submandibular gland. The amplicons were subcloned into the pCDNA3.1(+) mammalian expression plasmid and transformed into competent bacteria for plasmid DNA purification and DNA sequencing.
Alternatively spliced anoctamin1, anoctamin5, and anoctamin6
Sequencing of both strands of the cloned cDNA sequences, revealed evidence of multiple alternative splicing, especially for the anoctamin5 and anoctamin6 isoforms. The human anoctamin1 isoform is the best characterized with four alternatively spliced exons, designated a, b, c, d [10]. In the mouse submandibular salivary gland, DNA sequencing data from multiple subclones showed that the ac splice form was the primary anoctamin1 transcript. The calcium-dependent and voltage-dependent properties of the anoctamin1 channel vary depending on the anoctamin splice form [10]. In order to test whether anoctamin1 is alternatively spliced in the submandibular gland, six pairs of primers were designed to amplify overlapping portions of the 26 exons of the anoctamin1 gene products and analyzed on 2% agarose gel (Fig. 2). PCR amplification using primers for exon 2 and exon 9 showed a 500 bp main product band and an additional 570 bp band. Purification and DNA sequencing confirmed that the additional 70 bp sequence corresponds to anoctamin exon 7 (splice form b). No further evidence was found for the expression of other alternatively spliced anoctamin1 transcripts. Evidently, the mouse submandibular gland expresses two alternatively spliced isoforms: anoctamin 1ac and anoctamin1abc.
The sequencing analysis of anoctamin5 transcripts found an unexpectedly large number of alternatively spliced exons (Fig. 3). Using six primer pairs to amplify several exons of the anoctamin5 gene, PCR screening revealed multiple amplification products, especially when the primer pairs between exons 1-12, and exons 14-18 were used (Fig. 3B). A total of six alternatively spliced transcripts of anoctamin5 were detected: exon 5 (69 bp; anoctamin5a), which is thought to encode the cytosolic N-terminal part of the protein; exon 10 (135 bp; anoctamin5b), which codes for the first of eight transmembrane segment (TM1); exon 16 (108 bp; anoctamin5c), which codes for TM5; exons 19 and 20 (385 bp; anoctamin5d), which is the largest splice form and putatively codes for TM6 and TM7; exons 21 and 22 (285 bp; anoctamin5e), which account for TM7 and TM8; and exon 21 (107 bp; anoctamin5f), which codes for TM8. Most alternatively spliced transcripts are expected to result in function-compromised anoctamin5 proteins due to frame-shift associated termination during translation (anoctamin5d and anoctamin5f) or due to the in-frame deletion of putative transmembrane segments (anoctamin5b, anoctamin5c, anoctamin5e). Considering the low expression level of anoctamin5 compared to anoctamin1 (Fig. 1), the physiological significance of so many alternative splice variants of anoctamin5 in submandibular gland function is unclear and intriguing.
Mouse anoctamin6 transcripts were also analyzed by sequencing cDNA clones and multiple-exon-spanning PCR (Fig. 4). These experiments showed that among the 20 total exons of anoctamin6, alternative splicing occurred in three exons in the submandibular salivary gland. The first splice variant has been previously reported to involve an insertion of the additional exon 1a (72 bp; anoctamin6a), omission of the ATG start codon, and translation from a non-canonical CTG start codon [11]. Anoctamin6b is the second splice form, which skips the 285 bp exon 5, leading to a shortened cytosolic N-terminus of the anoctamin6 protein. The third alternative splice form (anoctamin6c) skips the 203 bp exon 18, which contains the extracellular loop between TM7 and TM8; this alternative splice produces a frame-shift and generates a truncated form of the anoctamin6 protein.
Unlike the anoctamin1 splice-variants that primarily arise from skipping exons in the N-terminal, several anoctamin5 and anoctamin6 alternative splice variants are expected to delete one or more putative transmembrane segments, leading to major alteration in the topological architecture of the membrane protein and its function. The specific details of the alternative splice sites of anoctamin5 and anoctamin6 are summarized in Table 2.
DISCUSSION
The activation of calcium-activated chloride channel activity is the rate-limiting step in salivary fluid secretion [12]. The knockdown of anoctamin1 channels using small interfering RNA has been found to reduce saliva flow in mice, suggesting that its function is essential in the formation of salivary fluid in vivo [2,12]. It is not known whether the other nine members of the anoctamin gene family, which share 20%-60% amino acid sequence identity with anoctamin1, play an additional role in salivary gland function. The identification of the expression of members of anoctamin gene family in the salivary gland is a first step toward clarifying the biological roles of other anoctamin proteins in saliva generation.
In our study, we tested for the expression of all members of the anoctamin gene family in the mouse submandibular salivary gland by RT-PCR. The primers chosen were designed to amplify the entire coding region of all members of the anoctamin gene family. As expected, anoctamin1 mRNA was expressed abundantly, in the submandibular gland, as were the mRNAs of anoctamin5, anoctamin6 and anoctamin10 (Fig. 1). Whether the latter anoctamin isoforms function as ion channels is not clear. When expressed heterologously in mammalian epithelial cell lines, the anoctamin5 and anoctamin10 proteins generate a very tiny current [13], raising the question of whether these isoforms are ion channels in the plasma membranes and whether they contribute to the calcium-mediated chloride efflux that drives salivary fluid secretion. The calcium-dependent ion channel activity of anoctamin6 appears to require cytosolic calcium in the micromolar range; however, previous reports do not agree whether anoctamin6 proteins are anion channels [14], cation channels [15], or a component of Ca2+-dependent phospholipid scramblases that translocate specific phospholipids between the two leaflets of the plasma membrane [8,16]. In our study, the expression of full transcripts of anoctamin3, anoctamin4, and anoctamin9 occurred in the submandibular gland, albeit at very low levels (Fig. 1). It is also possible that the transcripts for these anoctamins were not from the epithelial cells involved in exocrine secretion, but rather from other tissue components of the submandibular gland such as the nerves, vascular smooth muscle, and the blood cells. Indeed, anoctamin3 shows particularly high expression in neuronal tissues where it modulates Na+-activated potassium channels [17].
Alternative splicing generates an additional level of complexity in anoctamin expression in the submandibular gland. Anoctamin1ac and anoctamin1abc were the two forms expressed (Fig. 2). Based on the sequencing of multiple clones, our study did not show the presence of the short d splice form of anoctamin1. These alternative splice sites affect the N-terminal cytosolic part of the anoctamin1 protein, which determine the calcium sensitivity and voltage-dependent effects of the chloride channel function [10,18,19]. For example, the presence of the 22-amino acid segment encoded by the b splice variant (exon 7) in anoctamin1abc is expected to produce a chloride efflux that is resistant to channel inactivation under high intracellular Ca2+ levels [18], which may persist during the continued muscarinic stimulation of salivary acinar cells.
Anoctamin6 proteins also showed several alternatively spliced forms in the submandibular gland. The C-terminally spliced anoctamin6c likely generates a functionally compromised protein, since skipping exon 18 would lead to a translational frame-shift and premature truncation (Table 2). The splicing of exon 5 in anoctamin6b, however, is an in-frame deletion of 95 amino acids of the anoctamin6 protein. Ca2+ sensitivity may also be altered in anoctamin6b, similar to the N-terminal splice variants of anoctamin1 [10]. It is also possible that protein-folding and/or plasma membrane targeting may be affected by N-terminal splice variants, as N-terminal mutations produce intracellularly-directed anoctamin proteins [20]. However, additional experiments are needed to reach a definitive conclusion about whether the shorter N-terminus of anoctamin6 affects the ion channel function and/or phospholipid scramblase activity.
It is interesting to note that among the anoctamin family members, the anoctamin5 gene produced the most splice variants in the mouse submandibular gland. Mutations in anoctamin5 have been definitively linked to various pathologies, including gnathodiphyseal dysplasia, proximal limb-girdle muscular dystrophy, and distal non-dysferlin Miyoshi myopathy [6,7,21], underscoring the critical cellular role of this protein. In the submandibular salivary gland, at least six splice variants were expressed albeit at very low levels compared to anoctamin1 and anoctamin6. The anoctamin5a transcript would produce protein with a shortened N-terminus, much like the splice variants of anoctamin1 and anoctamin6. However, five of the splice variants we detected are hypothesized to produce major changes in the membrane topology of the protein, through in-frame deletions of exons that encode highly conserved transmembrane segments or by generating a frame-shift deletion of exons leading to a truncated protein (Table 2). Downstream of the excised transmembrane segments, the intracellular and extracellular aspects of membrane proteins may be reversed, resulting in channel proteins that are no longer functional. It is possible that these transcripts offer an evolutionary advantage by regulating specific target genes and/or proteins through an as yet unelucidated interference mechanism that affects transcripts and/or proteins by direct-binding to alternatively spliced regions of the transcript.
In this study, we report that full-length cDNA sequences were reverse-transcribed from transcripts of anoctamin1, anoctamin3, anoctamin4, anoctamin5, anoctamin6, anoctamin9, anoctamin10 found in the mouse submandibular gland. Multiple alternatively spliced anoctamin transcripts were also expressed, especially of anoctamin5, but this patterna was also found in anoctamin6, in which case most of the splice variants would likely lead to non-functional proteins. It is intriguing to speculate about the biological roles are performed by these alternatively spliced transcripts that have an infinitesimal possibility of producing intact channel protein in the organism, especially when the anoctamin isoform in question has been shown to have clear associations with human pathologies. It is possible that the biological role of these multiple alternatively spliced isoforms might reflect the functions of non-exocrine cell types in the submandibular glandular tissue, such as myoepithelial, endothelial, nerve, or endothelial cells, or alternatively may stem from specific acinar and ductal cells along the salivary gland ducts that may play enigmatic roles in exocrine secretion. Collectively, however, our results indicate that the major anoctamin protein responsible for the Ca2+-activated chloride efflux of salivary exocrine secretion is likely the anoctamin1ac form.




 XML Download
XML Download