

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Dental plaque is a complex biofilm composed of a consortium of multiple species. For a long time, the removal of dental plaque has been a key step in attempts to overcome periodontal disease. There are various combinations of nearly 700 bacterial species that are postulated to colonize the surfaces of the oral cavity [1,2]. Oral biofilms have a biodiverse, heterogeneous, organized structure that changes over time [3]. The internal structure of the dental biofilm could be the result of inter- and intraspecies coaggregation and coadhesion interactions aided by specific adhesin-receptor molecules [4,5]. Biofilm-embedded bacteria are less susceptible to antimicrobial agents than are planktonic bacteria. The biofilm structure serves as a physical barrier that limits the diffusion of agents, thereby severely limiting the effect of antibacterial treatment compared with the efficacy in planktonic bacteria [6,7].
Because in vivo studies of natural dental biofilms are limited and highly variable, the interpretation of the results of these in vivo studies is difficult. Difficulties with in vivo studies of the dental biofilm and its complex, heterogeneous structures have led to the development of laboratory biofilm model systems [8]. Although laboratory models cannot completely reproduce the complexity of the oral environment, they have certain advantages. An in vitro dental biofilm model circumvents the ethical conflicts that arise in clinical studies and can be used to analyze a variety of important in vivo processes in a highly reproducible fashion. These models have contributed considerably to elucidating the role of microbial interactions in biofilm development and the effect and working mechanisms of periodontitis-preventing agents [9].
Bacteriological studies revealed that gram-positive bacteria are early colonizers, including Streptococcus gordonii, Streptococcus oralis, Streptococcus sanguinis and other viridans group streptococci. As gingival inflammation progresses, gram-negative bacteria emerge in dental biofilms, including Fusobacterium nucleatum, which plays a crucial role as a bridging microorganism between early and late colonizers, and Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia, late colonizers present as a portion of the climax community in the biofilms at sites that exhibit progressing periodontitis [2,10]. Most biofilms in their natural environments are likely to consist of a consortium of species that influence one another synergistically or antagonistically. However, there is little knowledge of their structure, characteristics (including community dynamics), or response to antimicrobial agents.
While single-species biofilms have been studied extensively, we know notably little concerning multispecies biofilms and their interactions. The purpose of this study was to develop and evaluate an in vitro multispecies dental biofilm model that aimed to mimic the environment of chronic periodontitis.
MATERIALS AND METHODS
Bacterial strains and culture conditions
S. gordonii KN1, F. nucleatum ATCC 23726, A. actinomycetemcomitans ATCC 33384, and P. gingivalis ATCC 33277 were obtained from the culture collection of the Department of Oral Microbiology, Gangneung-Wonju National University Dental College. All bacteria were grown in trypticase soy broth (Becton Dickinson and Company, Sparks, MD, USA) containing 1 mg/mL of yeast extract (Becton Dickinson and Company), 5 µg/mL of hemin (Sigma Chemical Co., St. Louis, MO, USA), and 1 µg/mL of menadione (Sigma Chemical Co.) under anaerobic conditions (Bactron Anaerobic Chamber, Sheldon Manufacturing Inc., Cornelius, OR, USA) with an atmosphere of 90% N2, 5% CO2, and 5% H2.
Preparation of biofilms
The bacteria were grown in 12-well plates with a glass slip (round, 12-mm diameter). For four different single-species biofilms, the concentration of bacteria (S. gordonii, F. nucleatum, A. actinomycetemcomitans, and P. gingivalis) was adjusted by spectrophotometry (OD660) after incubation for 24 hours and then standardized by dilution with phosphate buffered saline (PBS) (pH 7.4) to 1×107 colony forming unit (CFU)/mL each. The glass slips were placed in the wells of a 12-well tissue culture plate. Next, the 12 wells were filled with 4 mL of culture medium and 25 µL of single-species bacteria, and incubated in anaerobic conditions at 37℃ for up to 48 hours, with a supply of fresh medium. For multispecies biofilms, we used the same method as for a single-species biofilm. However, 25 µL of medium was pooled with 4 different single-species bacteria. The glass slip was completely submerged in a 12-well plate. The growth kinetics was evaluated by generating growth curves for each experiment in triplicate. Briefly, the glass slip with the biofilm was harvested in the specific intervals (5, 24, 29, and 48 hours) and gently washed with sterile PBS to remove nonadherent bacteria. Next, 1,000 µL of crystal violet solution was added to the glass slip for 1 minute. Excess stain was removed with the administration of 2 mL of sterile PBS three times. A total of 1,000 µL of 80% ethanol and 20% acetone were added into the well and mixed on a shaker for 10 minutes. Next, 100 µL of prepared solution was transferred into new 96-well plates, and the absorbance was measured at 595 nm (ELx 800TM Absorbance microplate reader, BioTek, Winooski, VT, USA) to count the bacteria attached on a glass slip.
Confocal laser scanning microscope
Before the confocal laser scanning microscope (CLSM) analysis, the glass slip was gently washed with 2 mL of sterile PBS to remove nonadherent bacteria. After that, the glass slip was carefully mounted on a slide glass. The live/dead BacLight bacterial viability kit (Molecular Probes, Eugene, OR, USA) was used to view live or active cells (fluorescent green) and dead or inactive cells (fluorescent red) simultaneously. A total of 3 µL of SYTO 9 and 3 µL of propidium iodide were diluted in 1 mL of distilled water for the live/dead stain. A total of 100 µL of the reagent mixture was added to each glass slip, and the glass slips were incubated at room temperature in the dark for 15 minutes. Then, the reagent was removed, and the glass slips were cautiously washed with distilled water and examined at a magnification of 400× through a CLSM (IX71, Olympus, Tokyo, Japan). The CLSM images were obtained at 2-µm intervals until no fluorescent signal was observed.
Scanning electron microscopy
The glass slips with single-species and multispecies biofilm after 24 hours incubation were observed by scanning electron microscopy (VP-FE-SEM, CarlZeiss, Oberkochen, Germany). The glass slips with single-species and multispecies biofilms were washed twice with PBS. Glass slips with attached bacteria were fixed in 2.5% glutaraldehyde in PBS (pH 7.4) for 1 hour at room temperature. The fixed samples were washed three times with PBS for 10 minutes and dehydrated for 30 minutes in a graded series of ethanol. After critical point drying, the samples were mounted on stubs, coated with gold, and observed with SEM. Glass slips with single- and multispecies biofilms were observed by SEM (×10,000, ×30,000).
Determination of the minimal inhibitory concentration for planktonic and biofilm bacteria
Planktonic susceptibility tests of four different single-species and multispecies bacteria were performed by a broth microdilution assay according to the Clinical and Laboratory Standards Institute M07-A9 (2012) [11]. Antimicrobial agents were prepared with doxycycline (Sigma Chemical Co.) and chlorhexidine (Daewoongpharm Seoul, Korea). Overnight cultures of each bacterium were adjusted to 1×107 CFU/mL with bacterial culture broth, and 100 µL of broth was added to 96-well plates containing 1 mg/mL of doxycycline or 1,110 µM of chlorhexidine, and serial two-fold dilutions were prepared from working solutions. The bacteria were inoculated into serially diluted doxycycline or chlorhexidine in 96-well round bottom microtitration plates for final concentrations of 5×105 CFU/mL. The final volume was 100 µL in a microtitration plate well. The plates were read for turbidity after 18 hours for streptococci and 48 hours for F. nucleatum, A. actinomycetemcomitans, and P. gingivalis.
The biofilms were grown in 12-well plates with a glass slip for 24 hours. The glass slips with biofilm were transferred into new 48-well plates after washing with PBS. Approximately 1 mg/mL of doxycycline or 1,110 µM of chlorhexidine and serial two-fold dilutions from working solutions were added. The MIC was determined after 48 hours under anaerobic conditions. The MIC of the antibiotics was defined as the lowest concentration without turbidity of the broth. The tests were repeated at least twice.
RESULTS
Biofilm growth curves
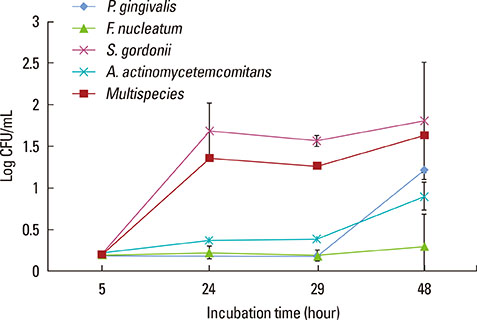
Fig. 1 represents the bacterial count of the biofilms over time. Comparison of the OD obtained at 595 nm indicates that S. gordonii and multispecies bacteria counts reached the stationary phase after 24 hours. S. gordonii, which is an early colonizer, and multispecies bacteria showed rapid growth patterns until 24 hours. F. nucleatum, the intermediate colonizer, showed a steady state until 48 hours. P. gingivalis and A. actinomycetemcomitans, which are late colonizers, showed a slow increase after 29 hours.
Structural analysis of biofilms
Each biofilm was collected at 24 hours and analyzed by CLSM and SEM. Fig. 2 represents confocal micrographs describing two-dimensional maximum projection images of the biofilms. Fig. 2 shows the vitality and thickness of biofilms. Multispecies biofilms were thicker than single-species bacteria. S. gordonii and multispecies biofilms showed a higher vitality than other species.
Fig. 3 represents SEM photographs of biofilms after 24 hours. F. nucleatum did not form a biofilm at 24 hours. Multispecies biofilms showed a multilayered structure compared to single-species biofilms. The CLSM and SEM examination revealed that the growth patterns of multispecies biofilms were similar to those of single-species biofilms. However, multispecies biofilms became thicker than the single-species biofilms, and networks between bacteria were formed.
Antibacterial activity
The MICs of bacteria used in this study are listed in Tables 1 and 2. In the multispecies model, the MIC value of planktonic bacteria for doxycycline was 0.0001 µg/mL, and the MIC value for the biofilm was 0.125 µg/mL. The MIC value of planktonic bacteria for chlorhexidine was 8.67 µM, and the MIC value for the biofilm was >1110 µM. The MICs of doxycycline and chlorhexidine were higher in the biofilm state than in planktonic bacteria. The MIC of doxycycline for the multispecies biofilm was higher than those for single-species biofilms of P. gingivalis, F. nucleatum, or A. actinomycetemcomitans. The MIC of chlorhexidine for the multispecies biofilm was higher than those for single-species biofilms of P. gingivalis or F. nucleatum.
DISCUSSION
Oral biofilms are a main focus of research in periodontal disease. In the present study, we attempted to develop an in vitro multispecies biofilm model that aimed to mimic the environment of chronic periodontitis, and we sought to evaluate the difference between single-species biofilms and multispecies biofilms using CLSM, SEM and MICs.
The bacteria used in this study are known periodontal pathogens, and they were composed of early, intermediate and late colonizers [10,12]. We observed a biofilm growth curve over time (Fig. 1). The multispecies biofilm seemed to enter the stationary phase after 24 hours; therefore, we selected 24-hour models in this study. However, the incubation period of a single-species biofilm with P. gingivalis, A. actinomycetemcomitans, and F. nucleatum might be necessary for longer than 48 hours. Considering previous studies, Prates et al. [13] used a 48-hour biofilm model of A. actinomycetemcomitans to investigate the bactericidal effect of photodynamic therapy with malachite green and red lasers, and Wright et al. [14] used a 72-hour biofilm model of P. gingivalis to compare the in vitro effect of metronidazole between the planktonic state and a single-species biofilm. However, we postulated the 24-hour multispecies biofilm model would minimize the possibility of contamination and increased the reproducibility.
In this study, we took CLSM and SEM images to observe the characteristics of biofilm structure. The CLSM and SEM examination revealed that the growth pattern of a multispecies biofilm was similar to those of single-species biofilms except F. nucleatum. With time, however, the multispecies biofilm became thicker than the single-species biofilms, and networks between bacteria were formed. This biofilm behavior was in agreement with previous studies [15,16]. Sanchez et al. [15] demonstrated that F. nucleatum was not detected in the subgingival biofilm model until 24 hours of incubation. P. gingivalis and A. actinomycetemcomitans could be measured inside the biofilm after 48 hours.
In the SEM image, the 24-hour biofilm model showed less adhesion of bacteria compared to previous studies. This study used a smooth glass slip to form biofilms. Conversely, in the previous studies, bacteria were grown on the disks with a rough surface such as ceramic calcium hydroxyapatite [15,17]. The difference in bacterial adhesion was most likely due to surface roughness and incubation time. However, it was postulated that the smooth glass slip was similar to the tooth surface in oral bacterial adhesion compared to disks with rough surfaces.
The MIC has been used as the gold standard for determining the antibacterial sensitivity for pathogenic bacteria [11,18]. Previous publications have reported that most bacteria embedded in various biofilms are far less sensitive to antibacterial treatment than are planktonic bacteria [7,14,19]. In this study, the MICs of doxycycline and chlorhexidine were observed to be higher in the biofilm state than in planktonic bacteria, as in previous studies (Tables 1 and 2). We observed that the antibiotic resistance in the biofilm state was increased approximately 100 to 10,000 times of resistance in planktonic bacteria regardless of the bacterial species. In the planktonic state, however, the antibiotic resistance of multispecies bacteria was higher than those of single-species bacteria by 10 to 100 times. In the biofilm state, we did not find differences in the antibiotic resistance between multispecies bacteria and S. gordonii bacteria. These results may be used in this experimental model, which is a 24-hour cultured biofilm model. S. gordonii, which is an early colonizer, showed a rapid growth pattern until 24 hours (Fig. 1). However, Figs. 2 and 3 showed that the multispecies biofilm became thicker than the S. gordonii biofilm, and networks between bacteria were formed.
This study is limited in that we did not clearly observe changes in the anaerobic bacteria inside the biofilm after 24 hours. However, the multispecies biofilm closely mimicked the condition of the oral cavity compared to single-species biofilms. The results obtained in this study suggested that a 24-hour multispecies biofilm has some advantages. First, it would minimize the possibility of contamination and increase reproducibility. Second, it would form a thicker biofilm and more complex networks than a single species of bacteria. Third, it showed a higher antibiotic resistance compared to a single-species biofilm. Therefore, a 24-hour multispecies biofilm can be easily applied in many experiments that use biofilms.
In conclusion, our results demonstrated that to mimic the natural dental biofilm, a multispecies biofilm composed of four bacterial species was grown. The 24-hour multispecies biofilm may be useful as a laboratory biofilm model system.





 XML Download
XML Download