

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Mesenchymal stem cells (MSCs) are multipotent progenitor cells that reside in the bone marrow and other tissues [1]. It is widely known that MSCs can differentiate into mesenchymal tissues like bone and cartilage under adequate conditions and they can be easily expanded in culture without losing their multipotency [2]. Many preclinical models for tissue engineering using MSCs have been reported and human clinical trials have been attempted recently.
MSCs can be used to regenerate damaged tissues when employed in specific scaffolds and implanted into defect sites. Although building the cell-scaffold composite with autologous stem cells would be preferred, there may be some patients whose MSCs are not available for tissue engineering due to lack of proliferation or differentiation potential. In that case, allogenic stem cells might be a reliable alternative. However, when allogenic cells are used, adjunctive immunosuppressants should be considered [3].
Immunosuppressants have provided great improvement in organ transplantation by suppressing the rejection of allografts, that is, graft-versus-host disease, which, in turn, has increased the survival rate of organ transplant patients. Cyclosporin A (CsA) and Tacrolimus hydrate (FK506) are widely-used, well-known immunosuppressants applied after kidney or heart transplantation [4,5]. CsA, a fungal metabolite, inhibits the production of T cell-derived soluble mediators such as interleukin (IL)-2, IL-3, and interferon (INF)-γ by inactivating calcineurin [6]. FK506 is a neutral macrolide with a different chemical structure from that of cyclosporine, but a mechanism of action similar to cyclosporine [7,8].
Some side effects of these immunosuppressive drugs have, however, been reported. Among those side effects, a considerable decrease in bone mineral density leading to osteoporosis has been demonstrated. Post-transplantation osteoporosis is a well documented phenomenon; patients treated with immunosuppressants often develop osteopenic conditions or bone fractures [9-11]. Similar high-turnover osteopenia related to systemic administration of immunosuppressants has also been observed in rats [12,13].
In contrast to in vivo findings, some in vitro studies have demonstrated that immunosuppressants may have osteogenic potential. Krocker et al. [14] investigated the effects of three immunosuppressants (FK506, CsA, and sirolimus) on cell proliferation and expression of a bone tissue-specific gene of human osteoblasts (MG63). They found that none of the examined drugs affected cell proliferation. Meanwhile, collagen III and XII, matrix metalloproteinase 2, Smad2, EGF receptor, annexin V, and osteonectin expression were increased by immunosuppressants, which implied a change in cellular differentiation.
Lee et al. [15] reported that rapamycin promoted the osteoblastic differentiation of human embryonic stem cells (hESCs). In their study, rapamycin induced the up-regulation of the early osteogenic markers, bone morphogenetic protein (BMP)-2 and Runx2, when hESCs were treated with rapamycin for 1 week. After continuous exposure of hESCs to rapamycin for 2 to 3 weeks, the mRNA levels of the late osteoblast differentiation marker genes, including osteocalcin, osteoprotegerin and osteonectin, were significantly elevated.
These conflicting in vivo and in vitro findings show that immunosuppressants may affect bone mineral metabolism in various ways that have not yet been clarified. Therefore, the effects of drugs on osteogenic differentiation of MSCs should be precisely analyzed prior to consideration of their clinical application for bone regeneration.
The purpose of this study was to investigate the effects of the immunosuppressants FK506 and CsA on proliferation and osteoblastic differentiation of rat MSCs in vitro.
MATERIALS AND METHODS
Cell isolation/preparation and culture
Rat MSCs were obtained from the femurs of 8-week-old Sprague-Dawley rats. The femurs were excised aseptically, their soft tissues were removed, and both ends of each femur were cut away. The marrow was flushed out by 20 mL of culture medium and the collected cell suspension was centrifuged at 400 g for 5 minutes. After the supernatants were removed, the pellet was released into 175 flasks with 20 mL culture medium. The cultures were incubated in a humidified atmosphere containing 5% CO2 at 37℃. The standard culture medium, which consisted of α-minimum essential medium, 15% fetal bovine serum, and antibiotics, was replaced after 24 hours to remove the nonadherent cells and again three times a week. At near confluence, the marrow cells were released using 0.1% trypsin and seeded in 24 96-well cell culture plates for subculture. The cultured cells in passage 2 to 5 were used for experiments and all biochemical assays were carried out at least in triplicate.
Immunosuppressants
FK506 (Cayman Chemical Co., Ann Arbor, MI, USA) and CsA (Cell Signaling Technology Inc., Danvers, MA, USA) were used from soluble sources. Each drug was diluted with phosphate buffered saline to concentrations of 50/500 nM.
Cell proliferation assay
Rat MSCs were seeded in a 96-well plate at 1×103 cells/well with the standard culture medium. Each immunosuppressant was added to each sample at various concentrations the next day. The culture medium and immunosuppressants were changed every other day. The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Lancaster, Morecambe, England) assay was used to determine the toxicity of immunosuppressants to stem cell proliferation at 1, 3, and 7 days. The medium was replaced with 200 µL fresh medium and 50 µL MTT solution was added. After 4 hours of incubation at 37℃, the MTT solution was removed and Dimethyl sulfoxide was added for the extraction of formazan. The optical density was measured with an automatic microplate reader (Thermo Max, Molecular Devices, Sunnyvale, CA, USA) at a wavelength of 540 nm.
Measurement of alkaline phosphatase (ALP) activity
The cells seeded in a 24-well plate at 5×103 cells/well were subcultured in 1 mL of the standard media supplemented with 10 mM β-glycerophosphate, L-ascorbic acid phosphate magnesium salt n-hydrate, and immunosuppressants. Each assay was performed in triplicate. The medium and drugs were refreshed three times a week. After 3, 7, and 14 days of subculture, the activity of ALP was examined using a SensoLyte pNPP ALP Assay kit (AnaSpec, Fremont, CA, USA). The culture media were removed and the cells were washed twice in 1× assay buffer. 300 µL of cell lysis buffer was added to each well and the cells were scraped. The cell lysates were centrifuged at 13,000 RPM for 10 minutes. p-nitrophenyl phosphate substrate was added to the supernatants (50 µL) and the reaction was allowed to occur at 37℃ for 30 minutes. Stop solution was then added and the absorbance was measured at 405 nm. The ALP activity of each well was expressed as micromoles of p-nitrophenol/30 min/µg. The ALP activity in each well was divided by the quantity of total protein in each well.
Mineralization nodule formation
Mineralization was assessed using alizarin red S staining. Rat MSCs were seeded in a 24-well plate at 5×103 cells/well. The cells were subcultured in 1 mL of the standard media containing 10 mM β-glycerophosphate, L-ascorbic acid phosphate magnesium salt n-hydrate, and immunosuppressants. At 21 days, the cells were fixed in 95% ethyl alchohol (EtOH), washed twice with distilled water, and stained for 5 minutes with Alizarin red S solution.
Real-time polymerase chain reaction (RT-PCR)
Total RNA extraction
Rat MSCs were plated at a density of 1×106 cells/60 mm petri dish. The next day was labeled "day 0", and the culture media were changed to media containing 10 mM β-glycerophosphate, L-ascorbic acid phosphate magnesium salt n-hydrate, and immunosuppressants.
RNA was extracted from the cultured cells on days 1 and 7. The procedures were summarized as follows: The cultured cells were lysed with easy-Blue (iNtRON Biotechnology Inc., Seongnam, Korea). Chloroform was added to the lysates to separate the phenol layer from the aqueous layer containing RNA. The upper aqueous layers were collected in a new tube and dehydrated with isopropanol. After washing with 75% EtOH, RNA pellets were air-dried and re-dissolved using diethylpyrocarbonate-treated distilled water.
RT-PCR
One µg total RNA was reverse transcribed for cDNA conversion using a SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. The sequences of the primers used for the RT-PCR are presented in Table 1.
The amplification reactions were performed in MicroAmp 96-well plates and each well contained SYBR Premix Ex Taq II, each PCR primer, ROX Reference Dye II, dH2O, and each template. The amplifications were carried out in a 7500 Fast RT-PCR System (Applied Biosystems, Foster City, CA, USA) and cycling conditions were as follows: initial denaturation step at 95℃ for 15 seconds, 40 3-step amplification cycles of 15 seconds denaturation at 95℃, 15 seconds annealing at 60℃, and 33 seconds extension at 72℃, dissociation stage. All RT-PCR reactions were run in duplicate.
Relative gene expressions were normalized to GAPDH expression and the data were presented as the fold change using the formula 2-ΔΔCT as recommended by the manufacturer (User Bulletin No.2(P/N 4303859), Applied Biosystems). When the value of 2-ΔΔCT is less than 1, the negative inverse of 2-ΔΔCT is presented as the fold change reduction in expression.
RESULTS
Analysis of cell proliferation
Cell proliferation assessment was done on days 1, 3, and 7. All groups showed evidence of increased cell proliferation over the whole examination period (Fig. 1). There were significant differences in optical dentity (OD) over time (P<0.05).
On day 1, there was no significant difference among any of the groups. However, the FK 506 groups showed a greater OD value than the control and CsA groups on days 3 and 7. No significant difference was found between the control and CsA groups.
The concentration of FK506 and CsA did not show any significant association with differences in cell proliferation.
Analysis of ALP activity
Fig. 2 shows the effects of immunosuppressants on the ALP activity of MSCs. ALP activity was assessed at 3, 7, and 14 days after treatment with immunosuppressants. On day 3, ALP activity of the control group was slightly greater than that of the other groups, but was not significantly different (P>0.05). The 50 nM and 500 nM FK506 (F50 and F500) groups showed higher ALP activity than the other groups on day 7 and the differences between the FK506 groups (F50 and F500) and 500 nM CsA group (C500) were significant (P<0.05). At a later point in time, the increase in the ALP activity level was most remarkable in the 50nM FK506 (F50) group and significantly different from the control and C500 group. The ALP activity of the CsA groups did not differ from the control group in any of the assessments (P>0.05).
Analysis of cultured cell mineralization
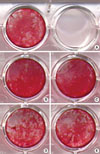
In the groups with FK506, Alizarin red S-(+) nodules were most prominent (Fig. 3). The control group showed more mineralization nodule formation than the CsA groups. In the experimental groups, the concentration of the drug made no difference in nodule formation macroscopically.
Analysis of the expression of mRNA for bone-related proteins
Quantifying the relative changes in gene expression was done on days 1 and 7. Immunosuppressants showed diverse outcomes depending on the drug concentrations, genes of interest, and time (Fig. 4).
Expression of osteopontin, an extracellular bone protein, was most markedly increased in the 500 nM FK506 (F50) group on day 1. A lower concentration (50 nM) also increased osteopontin mRNA expression over time. CsA decreased the expression of ostepontin relative to the control on day 1, but the expression then increased over time.
Treatment with FK506 had a clear positive effect on osteonectin mRNA expression on day 1, but the expression was diminished on day 7. Unlike FK506, CsA did not show a decrease in mRNA expression on day 7.
The fold changes of Runx2 mRNA expression were negative in all of the experimental groups on day 1, which implies that the immunosuppressants had an adverse effect on Runx2 expression. However, at later time points, the fold changes were reversed, which meant that Runx2 mRNA was expressed more than in the control group.
Type I collagen (Col-I), a mid-bone marker protein, showed increased expression when treated with 50 or 500 nM FK506 or 50 nM CsA. However, a higher concentration of CsA (500 nM) decreased Col-I expression during the experimental period.
Immunosuppressant treatment induced a down-regulation of Dlx5 gene expression on day 1, but on day 7, the expression was up-regulated compared to the control group.
DISCUSSION
The present study demonstrates that cell proliferation was promoted more in the FK506 groups than the control or CsA groups. ALP activity was significantly higher in the FK506 groups than the other groups. RT-PCR analysis showed elevated bone-related protein mRNA expression after FK506 treatment compared to the control group. These results indicate that FK506 may stimulate cell growth and also stimulate the differentiation of rat MSCs to osteoblasts. On the other hand, the effect of CsA on cell proliferation or differentiation was not evident.
Previously, several studies have reported the osteogenic effect of immunosuppressants using different cell types under different conditions. Isomoto et al. [16] assessed the effects of rapamycin, FK506, and CsA at 10 nM on the osteogenic differentiation of rat MSCs in vitro. Without dexamethasone (Dex), a well-known differentiation hormone, only FK506 showed an osteogenic effect on MSCs, which presented as higher ALP activity, calcium content, and osteocalcin content. With Dex, CsA and FK506 showed similar activity, even if it was less effective than in the control group (Dex only). Rapamycin did not have an osteogenic effect on MSCs, and they inhibited the osteogenic differentiation induced by Dex.
Kaihara et al. [17] used the local injection of BMP2-expressing recombinant adenoviral vector and FK506 into the calf muscle of rats and found that BMP2 mRNA expression was higher in the FK506 injection site than the control (only BMP2 viral vector injection site). In addition, ectopic bone formation was greater in the FK506 injection site.
Yoshikawa et al. [18] used the cultured bone construct in porous hydroxyapatite to test the effect of FK506 on the bone formation of implanted allogenic/isogenic bone construct in vivo. When the isografts were implanted subcutaneously, extensive bone formation was detected and the osteogenic parameters - ALP activity and osteocalcin content - were higher with FK506 than without FK506. In the allografts, high levels of osteogenic parameters and bone formation were found in the groups treated with FK506. Without FK506 administration, the allogenic bone constructs showed no bone formation or osteocalcin mRNA expression. Overall, the results of these recent studies suggest the potential of immunosuppressants, especially FK506, as an osteogenic supplement.
Kugimiya et al. [19] applied FK506 and CsA at a higher concentration (1,000 nM) to C17 and C2C12 cells for induction of osteogenic differentiation and demonstrated that FK506 was able to induce osteocalcin mRNA expression. However, when we examined the effect of FK506 and CsA on MSC proliferation at concentrations ranging from 10 to 1,000 nM, a high concentration of CsA had a negative effect on cell proliferation (data not shown). Based on those pilot study results, we chose lower concentrations than 1,000 nM. Even though the concentrations used in this study (50 and 500 nM) are higher than those for the systemic administration of transplant patients (FK506 at 0.01 µM, CsA at 0.1 µM), the results of MTT assay showed the positive effect of FK506 on MSC proliferation, while CsA did not affect cell proliferation. Further investigation into a more appropriate concentration is recommended.
The immunosuppressive action of FK506 and CsA is exerted through calcineurin inhibition. Cyclosporin-cyclophilin (immunophilin) complex inactivates calcineurin, resulting in the inhibition of cytokine expression including IL-2, 3, and INF-γ in T-cells. FK506 binds to the FK506-binding protein 12 (FKBP12) and the complex of FK506 and FKBP12 shows biological activity remarkably similar to that of cyclosporine [20]. However, FK506 and FKBP12 showed quite different effects on osteogenic differentiation in this study. This implies that the outstanding osteogenic effect of FK506 is not involved in calcineurin inhibition and BMP signaling is gaining attention as a possible pathway. Tang et al. [21] reported the effect of FK506, CsA, and rapamycin with or without BMP-4 on osteoblastic differentiation of mesenchymal cells: mouse osteoblastic-like cell line (MC3T3E1), mouse bone marrow stromal cell line (ST2), and mouse undifferentiated mesenchymal cell line (C3H10T1/2). Combined treatment with BMP and FK506 resulted in a significant increase in ALP activity in a dose/time dependent manner. FK506 also enhanced the positive effects of BMP on ALP and osteocalcin mRNA. However, osteocalcin expression was not affected by treatment with FK506 alone, without BMP. Kugimiya et al. [19] found that FK506 rapidly induced the phosphorylation of the BMP-dependent Smads, and the induction was blocked by Smad6; the activation of the Smads by FK506 is attenuated by FKBP12, and the binding of FKBP12 to the BMP receptors is suppressed by FK506. They suggested the hypothesis that the osteogenic signal induced by FK506 might be associated with the activation of the BMP receptors, probably by separating them from FKBP12.
In this study, most of the bone-related genes were expressed more in the FK506 groups than the control. Osteopontin, osteonectin, and Col-I genes were expressed more than in the control on day 1, but the intensity of expression had diminished by day 7. Runx2 and Dlx5 gene expression were reduced in the early stage, but at a later time, the levels of these genes were expressed more than in the control. A low concentration of CsA was also associated with an elevated level of osteonectin and a Col-I gene expression like FK506 on day 1, but a high concentration of CsA reduced the Runx2 and Dlx5 gene expression more severelyon day 1. Also, in 500nM CsA (C500) group, osteopontin and the Col-I gene were expressed less than in the control during the investigated period. On the other hand, osteocalcin genes were not detected in any group including the control group (data not shown). Osteocalcin is a bone-specific marker protein, which is generally a gene of interest in osteoblast differentiation studies. It has been reported that osteocalcin mRNA is not detected in less than 12 days during the proliferation and maturation stage, and was robustly expressed in the mineralization stage [22]. Osteocalcin gene expression is associated with the mineralization of osteoblasts and is considered a terminal marker of osteoblast differentiation. In this regard, our result that osteocalcin expression was not observed during the 7-day culture may be the result of relatively short-term exposure to the target drugs.
Recently, our group showed that ex vivo BMP-2 gene delivery using human gingival fibroblasts promotes bone regeneration in rats [23]. In the study, FK506 was successfully used as an immunosuppressant to diminish the immune response of rats to human cells. In addition, under FK506 coverage, considerable bone generation was observed even in the non-grafted control group. Therefore, although evidence on whether FK506 increases bone healing is still conflicting, it might be beneficial for bone regeneration in allotransplantation and/or xenotransplantation cases.
In conclusion, our results suggest that FK506 stimulates the osteoblastic differentiation of rat MSCs based on ALP activity and bone-specific gene expression. Therefore, FK506 might improve the results of bone regeneration in xenogenic or allogenic stem cell transplantation to treat bone defects. In addition, FK506 would be preferred over CsA in such cases.




 XML Download
XML Download