

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
With the help of effective screening and treatment methods, mortality of cervical cancer has been greatly reduced. However, cervical cancer still is the second leading cause of cancer deaths in women worldwide [1]. The single most important factor in cervical oncogenesis is high oncogenic risk human papillomaviruses (HPVs). HPVs interfere with Rb and p53 function to make cervical epithelial cells cancer-prone state. However, only a small portion of HPV-infected patients develop cervical cancer and this necessitates further studies of other factors, including cancer-related genes at the molecular level. Among the genes studied so far, fibroblast growth factor receptor (FGFR) is found constitutively activated [2]. Since the introduction of genome-wide, high-throughput research modalities, many studies to find disease-specific markers were conducted. As a result, they proposed several genes or gene products to have prognostic significance. Among the targets, significance of syndecans is reported in a few solid tumors and blood cancers.
Syndecans are plasma membrane proteoglycans and their cytoplasmic domain is thought to interact with the actin cytoskeleton [3]. They act as co-receptors by binding fibroblast growth factors (FGFs) and presenting them to FGFRs [3]. There are 4 different kinds of syndecans in vertebrates [4]. Among syndecans, which are expressed in various types of cells, syndecan-1 (SDC1) is expressed in epithelia and plasma cells [5].
Several studies on the relationship between syndecan expression and cancers were conducted. SDC1 expressions in various tumors, including head and neck cancer [6], hepatocellular carcinoma [7], mesothelioma [8], and lung cancer [9], were decreased. However, in some malignant tumors, such as endometrial cancer [10], ovarian cancer [11] and pancreatic cancer [12], SDC1 expression is increased. Those studies reported quantitative changes of SDC1 expression and the nature of the changes is dependent on the organ that the tumor has occurred. The syndecan family of matrix receptors is known to involve integrin (β4)-dependent signaling in human squamous carcinoma cells [13].
Prognostic significance of SDC1 expression is evaluated in several human cancers. Gallbladder cancer with SDC1 expression showed more frequent lymph node metastasis [14].
In this study, we compared SDC1 expression with several pathologic parameters in cervical cancers to find any significant association. To find possible cause of changes in SDC1 expression, we measured copy numbers of SDC1 in cervical cancers using fluorescent in situ hybridization (FISH).
MATERIALS AND METHODS
1. Patients and tumor samples
This study included tumor tissues surgically resected from 121 patients who visited Seoul St. Mary's Hospital and were diagnosed with uterine cervical cancer between 1999 and 2003. Patients' age was between 28 and 77 (mean, 50) years old. Most of patients were in FIGO stage I (69 cases, 57.0%) and II (48 cases, 39.7%). Eighty-two cases (68%) of patients were treated with combination chemotherapy consisting of cisplatin and etoposide before operation. Tumor-specific survival data (median, 61 months; range, 0.5 to 151 months) was available. The disease relapsed in 17 patients (14%), and 20 patients (17%) died of the disease. Using the tissues, tissue microarray (TMA) blocks were constructed and used for immunohistochemical staining and interphase FISH. Human tissue acquisition and its use followed the Institutional Review Board-approved protocol (CUMC10U917) at the Catholic University of Korea School of Medicine.
2. Immunohistochemistry
Sections from TMA blocks were transferred to ProbeOn Plus slides (Fisher Scientific, Pittsburgh, PA, USA) and incubated for two hours in 56℃ chamber (Agilent Technologies, Santa Clara, CA, USA). The sections were deparaffinized in xylene 3 times and rehydrated through 100%, 90%, 80%, 70% ethanol and Tris-buffered saline (TBS, pH 7.4). For antigen retrieval, the tissues were immersed in 10 mM sodium citrate buffer (pH 6.0) and boiled in a microwave for 20 minutes. After treating the tissues with 3% H2O2 in phosphate buffered saline (PBS), the tissues were incubated with diluted (1:50) mouse monoclonal antibody to SDC1 (Abcam, Cambridge, UK) at 4℃ overnight. After incubating the tissue with biotinylated anti-mouse antibody (Abnova, Walnut, CA, USA), TSA HRP System (PerkinElmer, Waltham, MA, USA) was used to amplify signal intensity. For visualization, liquid DAB+substrate chromogen system (Dako, Glostrup, Denmark) was used. Immunoreactivity of SDC1 was classified according to the percentage of tumor cells showing cytoplasmic stain; strong, >50% of cells stained; weak, 10-50% of cells stained; negative, <10% of cells stained. Two pathologists analyzed the immunoreactivity independently.
3. Fluorescent FISH
To synthesize SDC1 FISH probe, we used BioPrime Array CGH Genomic Labeling Module (Invitrogen, Carlsbad, CA, USA). BAC clone (PRP11-202B22; Invitrogen) was used as template and Spectrum Orange-dUTP (Abbott Molecular, Abbott Park, IL, USA) was used to label the probe. Aquarius Satellite probe of chromosome 2 (Cytocell, Cambridge, UK) was purchased as the reference probe. Location of homemade SDC1 probe was confirmed in metaphase spread of normal peripheral mononuclear cells. Tissue processing and hybridization was done using Paraffin Pretreatment Kit I (Abbott Molecular) and ThermoBrite (Abbott Molecular). We followed the manufacturer's recommended FISH protocols. We counted the number of fluorescent spots in at least 100 nuclei in each case.
4. Statistical analysis
Chi-square test and Fisher-exact test was used to evaluate significance of SDC1 expression in terms of pathologic features. The numbers of target spots and reference spots in FISH were compared using the chi-square test. For survival analysis, Kaplan-Meier method was used. To test the difference between survival curves of different groups, we used the nonparametric log-rank test. We used Cox's multivariate proportional hazard model to determine the prognostic values of selected clinicopathologic parameters. We used R ver. 2.10 (R foundation, Vienna, Austria) for statistical calculations and production of graphs.
RESULTS
1. Relationship between SDC1 expression and pathologic parameters
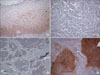
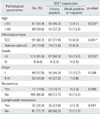
Normal squamous epithelial cells showed membranous pattern of SDC1 expression. Cytoplasm of normal cervical epithelial cells did not show SDC1 expression. On the contrary, cytoplasmic pattern of SDC1 expression was found in cancer cells in 101 cases (83.5%) of cervical cancers (Fig. 1). We compared the status of cytoplasmic SDC1 expression and some pathologic parameters. SDC1 was more frequently expressed in squamous cell carcinomas than adenocarcinomas and adenosquamous carcinomas (p=0.001). High grade tumors had less percentage of SDC1 expression than low grade tumors (p=0.036). We could not find any correlation between status of recurrence, lymph node metastasis, tumor stage and SDC1 expression (Table 1). Multivariate analysis showed that tumor grade and strong SDC1 expression were independent prognostic factors for survival (Table 2).
2. Copy number status of SDC1
There was no significant difference between the numbers of target (SDC1) spots and references spots (p>0.05) in the cases showing increased SDC1 immunoreactivity (Fig. 2).
3. Survival analysis
We analyzed patient survival among the groups with different SDC1 expression. Difference between the group of strong SDC1 expression and other groups was statistically significant (p=0.0219) (Fig. 3).
DISCUSSION
In this study, we showed that SDC1 expression is increased in cervical cancer and that most of the SDC1 accumulates in the cytoplasm of cancer cells. The finding that high grade tumors show decreased expression of SDC1 is consistent with a previous report [15].
Silencing SDC1 expression causes reduced focal adhesion plaque formation and enhanced cell spreading and motility on collagen I substrates [16]. Considering the fact that spreading and motility of cancer cells are highly related to invasion and distant metastasis, SDC1 positive tumors might have a more favorable biological behavior. This possible benefit of SDC1 may explain the findings that SDC1 expression is more common in low grade tumors and that patients with strong SDC1 expression have more favorable outcome in survival analysis.
We also showed that increased amounts of SDC1 is confined to the cytoplasm of tumor cells. Normal membranous distribution of SDC1 vanished in the neoplastic cells. Due to transposition of SDC1 from the cytoplasmic membrane to cytoplasm, the cells lose effective SDC1 on their surfaces. Considering the function of SDC1, these cells might lose connection to the extracellular matrix and move more freely than normal cells. Increased mobility of the cells can contribute to invasion and metastasis. Alteration of SDC1 distribution may be caused by degradation of the glycosaminoglycan chains, or expression of a mutated core protein that cannot undergo glycanation [17].
In one report, an inverse correlation between the expression of SDC1 in the tumor and lymph node metastasis was suggested [18]. However, they failed to show statistical significance, as we did.
In other malignancies, such as endometrial cancers [19] and invasive ductal breast carcinomas [20], SDC1 expression is reported to be associated with better prognosis. However certain malignancies, such as gallbladder cancer [14] and prostate cancer [21], it has been reported to have shorter survival time with positive SDC1 expression. This contradictory effect of SDC1 might be due to the difference of the site of origin that the tumors have come from.
SDC1 can be detached from the cytoplasmic membrane and become soluble form. The soluble SDC1 is associated with shorter survival in chronic lymphocytic leukemia [22] and lung cancer [23]. SDC1 expression of the bone marrow environment is associated with shorter event-free survival of multiple myeloma [24]. Adverse effect of SDC1 expression in tumor stroma has also been reported in oral carcinomas [25] and pancreatic cancer [26].
Decreased expression of SDC1 can be ascribed to increased degradation of the protein in cancer cells without any change in the gene expression. As a mechanism of decreased SDC1, decreased biosynthesis of the protein is suggested in gastric cancer [27]. Sometimes changes in copy number of a gene result in alterations in gene expression. To find a cause of SDC1 expression change, we screened the copy number of SDC1 and found no alteration in its copy number. It seems that alteration of SDC1 expression is caused by changes other than copy number alteration. The mechanism of SDC1 expression in malignant cell is not known to date.
We found two reports claiming that SDC1 expression did not predict survival [28,29]. In those articles, authors scored immunoreactivity of SDC1 on the surface of cells. SDC1 can be found on the surface of normal epithelial cells, and cervical cancer cells may retain SDC1 expression on the surface. Because we focused on abnormal location of SDC1 in tumor cells, we scored cytoplasmic expression of SDC1. With this scoring scheme, we found that strong SDC1 expression was associated with better survival. This explains the difference of our results and previous reports.
In conclusion, we showed intense SDC1 expression is associated with better prognosis in cervical cancer. Tumor grade and histological type is also related to the SDC1 expression. The alteration of SDC1 expression in cervical cancer is not caused by copy number changes of SDC1 in the tumor.



 XML Download
XML Download