

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Gastric cancer is one of the most common malignant tumors worldwide, and its morbidity and mortality are still high. Generally, it is diagnosed at an advanced stage, leading to high metastasis and poor prognosis.Therefore, the identification of an effective therapeutic target that inhibits the proliferation and metastasis of human gastric carcinomas is necessary.
Suppressor of variegation, enhancer of zeste, trithorax (SET) and myeloid-nervy-deformed epidermal autoregulatory factor-1 (MYND) domain-containing protein 3 (SMYD3) is a member of the histone methyltransferases; it can specifically methylate histones at H3k4, H4K5, and H4K20 [12] and is involved in signal transduction [3]. Previous studies have demonstrated that SMYD3 is overexpressed in hepatocellular, colorectal, cervical, and breast cancer [45678]. Additionally, our recent study indicated that SMYD3 expression might be an independent predictive factor for overall survival in gastric cancer [9]. However, the direct role and detailed molecular mechanisms of SMYD3 in gastric cancer are still unclear.
Activated ataxia telangiectasia mutated (ATM) triggers the phosphorylation of its downstream targets p53 and checkpoint kinase 2 (CHK2), which phosphorylate Cdc25C at Ser 216, contributing to G2/M phase checkpoints [10]. Cdc25C, a cyclin-dependent kinase (CDK)-activating kinase, is necessary for the activation of CDK1 and further progression through the cell cycle. The cyclin-dependent kinase 1 (CDK1; Cdc2)/cyclin B complex plays a significant role in the regulation of the G2/M phase. Cdc25C is phosphorylated at Ser 216 in the active state and catalyzes cyclin B/CDK1, thereby allowing for the unscheduled activation of CDK-cyclins, which are associated with G2/M progression.
In this study, the effects of SMYD3 on the proliferation, migration, and invasion of gastric cancer cells were investigated. In contrast to previous results obtained for breast cancer, our present data indicated that the absence of SMYD3 could halt cells in G2/M phase via the ATM-CHK2/p53-Cdc25C pathway. Furthermore, the rates of gastric cancer cell migration and invasion were reduced by knocking down SMYD3. In contrast, the overexpression of SMYD3 could promote the proliferation, migration, and invasion of GES-1 normal gastric mucosal cells. These findings indicate that SMYD3 might be a useful therapeutic target in human gastric carcinomas, and the ATM signaling pathway might be involved in the SMYD3-mediated regulation of proliferation and migration in gastric cancer cells.
MATERIALS AND METHODS
Cell lines and transfection
MGC-803 and AGS human gastric cancer cells were cultured in DMEM (Gibco, Grand Island, NY, USA) and human normal gastric mucosal cells GES-1 were cultured in RPMI 1640 medium (Gibco) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin G, and 0.1 mg/mL streptomycin. Cultures were maintained in a 5% CO2 humidified atmosphere at 37°C. The plasmid pcDNA5-TO/TAP-DEST-SMYD3 was a gift from Professor Philip Tucker (Institute for Cellular and Molecular Biology, University of Texas, Austin, TX, USA) [2]. siRNAs were synthesized by Ribo Biological Company (Shanghai, China) and their sequences were as follows: siRNA targeting SMYD3 (si-SMYD3), sense strand, 5′-CAAGGAUGCUGAUAUGCUAdTdT-3′; antisense strand, 3′-dTdT GUUCCUACGACUAUACGAU-5′. The sequence of the control siRNA (si-control) was confidential, i.e., it was not provided by the manufacturer. Transient transfection was performed using TurboFect reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA), according to the manufacturer's instructions.
RNA extraction, cDNA synthesis, and reverse transcription quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from gastric cells using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) under RNase-free conditions. After quantification using a photometer, complementary DNA was synthesized by M-MLV reverse transcriptase with 2 μg of total RNA. RT-qPCR was performed using the SYBR Green Supermix obtained from DBI (DBI-2043; Delaware Biotechnology Institute, Newark, DE, USA) and detected using the ABI Step One system (Thermo Fisher Scientific, Inc.). Primers were obtained from Invitrogen (Beijing, China) and their sequences are shown in Table 1. Real-time polymerase chain reaction (PCR) was performed as follows: 95°C for 2 minutes, followed by 40 cycles of 95°C for 10 seconds, 65°C for 30 seconds, and 72°C for 30 seconds. The 2−ΔΔCT method was used to calculate relative transcription levels. All experiments were repeated at least 3 times.
Table 1
Specific primers used in this study

ATM = ataxia telangiectasia mutated; CHK2 = checkpoint kinase 2; CDK1 = cyclin-dependent kinase 1; SMYD3 = suppressor of variegation, enhancer of zeste, trithorax and myeloid-nervy-deformed epidermal autoregulatory factor-1 domain-containing protein 3; GAPDH = glyceraldehyde 3-phosphate dehydrogenase.
Western blotting
After transfection for 48 hours, cells were washed with ice-cold phosphate-buffered saline (PBS) twice and lysed at 4°C with lysis buffer containing protease inhibitors and protein phosphatase inhibitor (Solarbio, Beijing, China) for 30 minutes. The lysates were centrifuged at 10,000 rpm for 10 minutes and the supernatants were mixed with sodium dodecyl sulfate (SDS) sample buffer. The soluble proteins were separated by SDS-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. The membranes were blocked in blocking buffer (5% not-fat dry milk in PBS) for 1.5 hours at 20°C, and then blotted with appropriate primary antibodies against ATM (rabbit anti human monoclonal antibody; dilution, 1:1,000; ab32420; Abcam, Cambridge, MA, USA), ATM (phospho S1981) (rabbit anti human monoclonal antibody; dilution, 1:1,000; ab81292; Abcam), CHK2 (rabbit anti-human polyclonal antibody; dilution, 1:500; bs-1391R; Bioss, Beijing, China), CDK1 (rabbit anti-human polyclonal antibody; dilution, 1:500; bs-1341R; Bioss), CyclinB (rabbit anti-human monoclonal antibody; dilution, 1:500; SRP10402; Saierbio, Tianjin, China), p53 (mouse anti human polyclonal antibody; dilution, 1:1,000; sc-126; Santa Cruz Biotechnology, Santa Cruz, CA, USA), p21 (mouse anti human polyclonal antibody; dilution, 1:1,000; sc-6246; Santa Cruz Biotechnology), Cdc25C (rabbit anti-human monoclonal antibody; dilution, 1:1,000; ab32444; Abcam), Cdc25C (phospho S216) (rabbit anti human monoclonal antibody; dilution, 1:1,000; ab32051; Abcam), SMYD3 (rabbit anti human monoclonal antibody; dilution, 1:1,000; ab187149; Abcam), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (mouse anti human; dilution, 1:5,000; UM4002; Utibody, Beijing, China). Subsequently, infrared-stained goat anti-rabbit (IRDyew680RD; 926-68071; LI-COR Biotechnology, Lincoln, NE, USA) and goat anti-mouse (IRDyew800CW; 926-32210; LI-COR Biotechnology) secondary antibodies were incubated with the membrane at a dilution of 1:10,000. The signals were then visualized using the Odyssey Imaging System (LI-COR Biotechnology).
Migration and invasion analysis
For the wound healing assay (2D migration assay), cells were seeded in 6-cm dishes and transfected with siRNAs or plasmids according to the manufacturer's instructions. Six hours later, a wound was made and images were captured immediately. The cells were continuously incubated with serum-free medium. The wound was photographed every 6 hours, and the migration rates were calculated based on the width of the wounds.
For the Transwell assay (3D migration assay), Transwell chambers (Corning Life Sciences, Lowell, MA, USA) were used. Cells were seeded in the upper chamber with serum-free medium after transfection, and the bottom chambers were filled with DMEM containing 10% FBS. After 16 hours of incubation at 37°C, the non-migrated cells on the upper membrane surfaces were removed and the migrated cells on the lower surface of the upper chamber were fixed with methanol, washed with PBS thrice, and then stained with DAPI (Solarbio) for 1 hour in the dark. Cell migration was quantified by counting cells using a fluorescence microscope. In addition, for invasion assays, cells were seeded in a Matrigel-coated Transwell (BD Biosciences, Franklin Lakes, NJ, USA), and viable cells were quantitated after 24 hours using crystal violet (Solarbio).
3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay
Cells were seeded in 96-well plates at a density of 5×104 cells per milliliter. After starvation in serum-free medium for 24 hours, cells were transfected with siRNAs or plasmid. MTT (Solarbio) was added to each well at 24, 48, or 72 hours after the treatment, followed by incubation at 37°C for 4 hours. Subsequently, all medium was removed and 150 μL per well of dimethyl sulfoxide was added to dissolve formazan. The absorbance value at a wavelength of 570 nm was read using a Synergy4 microplate reader (BioTek USA, Winooski, VT, USA). Cell viability was calculated as a percentage relative to untreated cells according to the following equation:
where OD means optical density.
Each sample was examined in duplicate and the experiment was repeated 3 times.
Clonogenic assay
After transfection with siRNA or plasmid for 24 hours, the cells were seeded in a 6-well plate at a density of 1,000 cells per well. Subsequently, the cells were cultured for 10 days to obtain visible colonies. After they were washed with PBS twice, cells were fixed with methanol for 15 minutes at room temperature and stained with Giemsa for 30 minutes. Colonies comprising at least 50 cells were then counted.
Cell cycle detection
MGC-803 or AGS cells were seeded in 6-well plates at a density of 4×104 cells per well and transfected with si-control or si-SMYD3 RNAs. Twenty-four hours later, cells were digested with trypsin, collected by centrifugation, washed with PBS twice, and dropped in ice-cold 70% ethanol for 12 hours at −20°C. Subsequently, cells were centrifugated, washed with PBS twice, and then incubated in PBS containing 50 μg/mL propidium iodide (PI; Solarbio) and 0.1% RNase A (Solarbio) at 4°C in the dark. Stained cells were analyzed using the Accuri C6 flow cytometer (Accuri, Ann Arbor, MI, USA), and counted with at least 10,000 events. Data were analyzed using ModFIT (Verity Software House, Inc., Topsham, ME, USA).
Statistical analysis
Data are summarized as means±standard deviation (SD). Statistically significant differences were evaluated by Student's t-tests (P<0.05). Statistical analyses were performed using GraphPad Prism version 5.00 (GraphPad, San Diego, CA, USA). All experiments were carried out at least 3 times.
RESULTS
SMYD3 is essential for the migration capability of gastric cells
To explore whether SMYD3 is involved in the migration of human gastric cancer cells, RNAi technology was used to knock down the endogenous expression of SMYD3 in MGC-803 and AGS cells. As shown in Fig. 1D, cells transfected with si-SMYD3 exhibited a notable reduction in SMYD3 expression for 10 days compared with expression in control cells. Subsequently, the effect of SMYD3 on cell migration was evaluated by a wound healing migration assay (2D migration assay) and a Transwell assay (3D migration assay). Fig. 1A summarizes the differences between the si-SMYD3 treatment group and the control group in the wound healing migration assay. In the control group, cells converged at 72 hours but not SMYD3 group. Similarly, the results of the 3D migration assay confirmed that the siRNA-mediated suppression of SMYD3 could significantly lower the migration rate (Fig. 1B). In addition, we investigated the effects of SMYD3 on the invasive activity of gastric cancer cells in Matrigel-coated Transwells. As shown in Fig. 1C, the invasive ability of MGC-803 cells or AGS cells was also suppressed by the knock-down of SMYD3.
Fig. 1
Effect of SMYD3 on the migration and invasion of gastric cells. After transfection with SMYD3-specific siRNAs or an overexpression plasmid, the migration ability of cells was detected by a wound healing assay (A) and a Transwell assay (B), and the invasion ability was analyzed by a Matrigel-coated Transwell assay (C). The interference efficiency of SMYD3 at various time points was detected by RT-qPCR (D).
SMYD3 = suppressor of variegation, enhancer of zeste, trithorax and myeloid-nervy-deformed epidermal autoregulatory factor-1 domain-containing protein 3; RT-qPCR = reverse transcription quantitative polymerase chain reaction; si-SMYD3 = siRNA targeting SMYD3.
*P<0.05; †P<0.01; ‡P<0.001 vs. control cells.
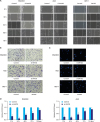
Furthermore, the same analyses were performed using GES-1 normal gastric mucosal cells after transfection with the SMYD3 overexpression plasmid (pcDNA5-TO/TAP-DEST-SMYD3). The overexpression of SMYD3 promoted the migration and invasion of gastric cells (Fig. 1). Taken together, these results suggested that SMYD3 was essential for the migration and invasion of gastric cancer cells.
Knock-down of SMYD3 could decrease cell viability in gastric cancer cells
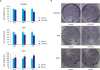
To explore the effect of SMYD3 on the proliferation of gastric cells, an MTT assay was performed. The knockdown of SMYD3 by siRNAs inhibited the proliferation of MGC-803 and AGS gastric cancer cells in a time-dependent manner (Fig. 2A). Furthermore, a clonogenic assay was performed to verify the results obtained from the MTT assay. As shown in Fig. 2B, markedly fewer clones were observed in the si-SMYD3 treatment group than in the control group, suggesting that SMYD3 plays important roles in the proliferation of human gastric cancer cells. To verify this hypothesis, an MTT assay and clonogenic assay were further performed using GES-1 gastric cells, characterized by a low level of endogenous SMYD3. As shown in Fig. 2, cells transfected with SMYD3 exhibited greater viability and clonogenic potential than those of control cells, indicating that SMYD3 could improve the proliferation of gastric cells.
Fig. 2
Knockdown of SMYD3 could inhibit the proliferation of gastric cancer cells. MGC-803 and AGS gastric cancer cells were transfected with si-SMYD3 and cell viability was analyzed by MTT assays at various time points (A). A clonogenic assay was performed after transfection for 10 days (B). The same experiments were performed using GES-1 normal gastric cells transfected with a control vector or SMYD3 overexpression vector.
SMYD3 = suppressor of variegation, enhancer of zeste, trithorax and myeloid-nervy-deformed epidermal autoregulatory factor-1 domain-containing protein 3; MTT = 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide; si-SMYD3 = siRNA targeting SMYD3.
*P<0.05; †P<0.01; ‡P<0.001 vs. control cells.
SMYD3 knockout induced G2/M arrest via the ATM-CHK2/p53-Cdc25c pathway
To further determine the role of SMYD3 in the proliferation of gastric cancer cells, cell cycle distribution analyses were conducted by flow cytometry using MGC-803 and AGS cell lines. The anti-proliferation effect of the siRNA-mediated suppression of SMYD3 might occur by G2/M cell cycle arrest. Fig. 3A depicts a representative profile of the cell cycle distribution in gastric cancer cells. In total, 14.01% of MGC-803 cells treated with si-control RNAs were in G2/M phase. si-SMYD3-transfected cells exhibited a substantial increase in the G2/M fraction; approximately 23.92% of cells were arrested at the G2/M phase of the cell cycle when endogenous SMYD3 was knocked down by siRNAs (Fig. 3B). For AGS cells treated with si-control RNAs, 13.50% were in G2/M phase, whereas approximately 26.88% of cells were arrested at the G2/M phase of the cell cycle when endogenous SMYD3 was knocked down by siRNAs.
Fig. 3
The anti-proliferation effect of SMYD3-targeted siRNA might be due to G2/M cell cycle arrest. MGC-803 and AGS gastric cancer cells were transfected with si-SMYD3, and cell cycle progression was evaluated by flow cytometry after 24 hours (A); the cell cycle distribution was further analyzed (B).
SMYD3 = suppressor of variegation, enhancer of zeste, trithorax and myeloid-nervy-deformed epidermal autoregulatory factor-1 domain-containing protein 3; si-SMYD3 = siRNA targeting SMYD3.
*P<0.05; †P<0.01; ‡P<0.001 vs. control cells.

To identify the molecular mechanisms by which SMYD3-targeted siRNA induced G2-phase arrest, the effects of the suppression of SMYD3 on the expression of key cell cycle regulators were investigated by quantitative real-time PCR and western blotting. As shown in Fig. 4A and 4B, when endogenous SMYD3 was suppressed by siRNAs for 24 hours, the expression levels of ATM (both total ATM and phosphorylated ATM), p53, CHK2, and p21 increased, whereas CDK1 and cyclin B, downstream genes of the ATM pathway and key regulators in G2 phase, were downregulated. These results suggested that the absence of SMYD3 could activate the ATM-CHK2/p53 pathway and stimulate the phosphorylation of Cdc25C, thereby inducing arrest at the G2/M checkpoint.
Fig. 4
The absence of SMYD3 could activate the ATM- CHK2/p53 pathway and stimulate the phosphorylation of Cdc25C. The mRNA levels of ATM, CHK2, Cdc25C, Cyclin B, CDK1, and p53 were determined by real-time qPCR in MGC-803 cells and AGS cells after 24 hours (A). The expression level of ATM and proteins associated with G2/M phase were further evaluated by western blotting after 48 hours (B). GAPDH was used as an internal control.
SMYD3 = suppressor of variegation, enhancer of zeste, trithorax and myeloid-nervy-deformed epidermal autoregulatory factor-1 domain-containing protein 3; ATM = ataxia telangiectasia mutated; CHK2 = checkpoint kinase 2; CDK1 = cyclin-dependent kinase 1; qPCR = quantitative polymerase chain reaction; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; si-SMYD3 = siRNA targeting SMYD3.
*P<0.05.

DISCUSSION
SMYD3 possesses histone methyltransferase activity; in particular, it catalyzes the methylation of histones at H3K4, K4K5, and H4K20, and elicits oncogenic effects by activating the transcription of downstream target genes in hepatocellular, colorectal, cervical, and breast cancer [567811]. Recently, we obtained the first evidence that the overexpression of SMYD3 might also be a risk factor in the biological behavior and prognosis of gastric cancer [9]. However, the direct role and detailed molecular mechanisms of SMYD3 in gastric cancer are unclear. In the present study, we demonstrated that a lack of SMYD3 could suppress migration, proliferation, and cell cycle progression in gastric cancer cells, confirming that SMYD3 might be a useful therapeutic target in human gastric carcinomas.
A previous study showed that the knockdown of SMYD3 induces G0/G1-phase arrest in MDA-MB-231 human breast cancer cells [12], and we verified this result in the human breast cancer cell lines MCF-7 and T47D. However, in MGC-803 gastric cancer cells, we demonstrated that the down-regulation of SMYD3 induced G2/M-phase arrest, but not G0/G1-phase arrest, indicating that SMYD3 might have different functions in different types of carcinomas. Furthermore, our results showed that the activation of the ATM-CHK2/p53-Cdc25C pathway might play an important role in the G2/M-phase arrest induced by SMYD3 suppression. ATM is a critical regulator of the DNA damage response and cell cycle control. Following its activation, downstream effectors, such as CHK2 and p53, are phosphorylated and activated, leading to further transmission of checkpoint signals. Interactions between CDKs and cyclins promote cell cycle progression. CDKs are activated by dephosphorylation at Thr-14 and Tyr-15 of Cdc25, which complete phase transitions, including CDK1, a regulator of mitotic entry [13]. Cdc25 can be phosphorylated by CHK2 on Ser-323 (Cdc25B) and Ser-216 (Cdc25C), and compared with Cdc25A and Cdc25B, Cdc25C is a much more representative marker in G2/M regulation. Previous studies have found that Cdc25A plays an important role at the G1/S-phase transition, Cdc25B undergoes activation during S-phase, and activated Cdc25C leads to positive feedback loops and stimulates cells entry into mitosis [14]. Interestingly, Sun and colleagues have found that spSet1p, a histone H3K4 methyltransferase, is involved in the regulation of the ATM pathway in yeast [15]. Similarly, our data showed that SMYD3, a SET domain-containing histone H3K4 methyltransferase, might modulate an ATM-related pathway and regulate G2/M progression in gastric cancer cells.
Increasing evidence suggests that the ATM pathway also plays essential roles in epithelial-mesenchymal transition (EMT), drug resistance, and cancer metastasis [15161718192021222324]. Sarris and colleagues [25] have reported that snail1 and other EMT markers are significantly upregulated in the livers or colons of wild-type mice after treatment with diethylnitrosamine or 1,2-dimethylhydrazine and dextran sodium sulfate. However, these phenomena were not observed in SMYD3-KO mice, indicating that SMYD3 might play an important role in the regulation of EMT during liver and colon carcinogenesis. Furthermore, their research showed that the mechanisms underlying the regulatory effects of SMYD3 on these target genes involves its potentiating effect on RNA pol-II-mediated transcription and histone methylation. However, related research focused on gastric cancer is lacking [12]. It is possible that SMYD3 also participates in the regulation of EMT and chemosensitivity, and this will be a focus of future research in our lab.
In summary, to the best of our knowledge, this study provides the first evidence that a lack of SMYD3 inhibits the migration and proliferation of gastric cancer cells and halts cells in G2/M phase via the ATM-CHK2/p53-Cdc25C pathway. Further studies of the molecular mechanisms underlying the effects of SMYD3 in various cancer cell types might provide a basis for the development of novel therapeutic approaches to human cancers.
 XML Download
XML Download