

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Infection with the Epstein-Barr virus (EBV), a ubiquitous oncogenic g-type herpes virus, is implicated in the etiology of several lymphoid and epithelial malignancies, including Burkitt's lymphoma, Hodgkin's lymphoma, nasal NK/T cell lymphoma, nasopharyngeal carcinoma, and a subset of gastric carcinomas (GCs).123 EBV-associated GC (EBVaGC) is defined as the monoclonal proliferation of carcinoma cells with latent EBV infection, which can be demonstrated by in situ hybridization targeted at EBV-encoded small RNA (EBER). In EBVaGC, virtually all cancer cells contain EBV DNA sequences, and the EBV terminal repeat sequences have uniform lengths, implying that the tumor can arise from a single EBV-infected cell and that the EBV genome persists during malignant transformation and proliferation.45 About 10% (1.3% to 20.1%) of GCs are EBV-positive;678 however, the prognostic effect of EBV positivity in GC is controversial. While some studies have shown significantly better prognosis in EBVaGC than in EBV-negative GC (EBVnGC),9101112 other studies failed to observe this prognostic difference.13141516 Nonetheless, a recent international large-scale meta-analysis of 4,599 cases revealed that patients with EBVaGC had longer survival than those with EBVnGC.17
Recently, The Cancer Genome Atlas (TCGA) research network proposed a new molecular classification, according to which GC is divided into four subtypes: EBV-positive tumors, microsatellite-instable tumors, genomically stable tumors, and tumors with chromosomal instability. EBV-positive tumors display recurrent PIK3CA mutations, extreme DNA hypermethylation, and enhanced expression of JAK2, CD274 (also known as programmed death receptor-ligand [PD-L] 1), and PDCD1LG2 (also known as PD-L2).18
Histologically, EBVaGC is characterized by marked intra- or peritumoral immune cell infiltration. In our previous study, we classified EBVaGC into three histological subtypes according to the cellular immune responses that affect prognosis: lymphoepithelioma-like carcinoma (LELC), carcinoma with Crohn's disease-like lymphoid reaction (CLR), and conventional adenocarcinoma (CA).19 The prognostic value of immune reactions in EBVaGC suggests that tumor microenvironment is a very important factor in the progression of EBVaGC.
Numerous studies have focused on the relationship between host cellular immune responses and EBVaGC prognosis. In this article, we reviewed the clinicopathological features of EBVaGC and EBV-associated immune responses in patients with EBVaGC.
Clinicopathological and Histological Features of Epstein-Barr Virus-Associated Gastric Carcinoma
1. Clinicopathological characteristics
According to the meta-analysis of 15,952 cases, EBV-positive GC occurs more frequently in male than in female patients. EBV-positive tumors usually arise in the cardia or the body of the stomach rather than in the antrum. Tumors in the post-surgical gastric stump/remnants are four times more likely to be EBV-positive than other GCs.20 There was no significant correlation between age and EBV positivity.16192021 Several studies have found an apparently higher incidence of synchronous multiple carcinomas in EBVaGC than in EBVnGC.222324 Smoking is one of the risk factors of EBVaGC.25 The influence of Helicobacter pylori infection on EBVaGC emergence is controversial.721262728 The presence of EBV-positive tumors negatively correlates with the TNM stage parameter as well as with the values of its individual components (primary tumor site, regional lymph node involvement, or presence of distant metastasis).17 Overall, EBV positivity is associated with favorable prognosis.9101112
2. Histological characteristics
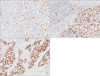
In histological examinations, marked intra- or peritumoral immune cell infiltration is usually detected in EBVaGC samples. Previously, we divided EBVaGC into three histological subtypes according to the microscopic characterization of host cellular immune responses: LELC, CLR, and CA. Typical LELC was defined by (1) a well-defined tumor margin, (2) dense lymphocytic infiltration when the number of tumor-infiltrating lymphocytes (TIL) was greater than that of tumor cells, (3) indistinct cytoplasmic borders and a syncytial growth pattern with poorly formed glandular structures, and (4) absence of desmoplasia (Fig. 1A). CLR was characterized by (1) patchy lymphocytic infiltration with three or more lymphoid follicles with active germinal centers per tissue section at the advancing edge of the tumor, (2) lower number of lymphocytes compared to tumor cells, (3) frequent tubule or gland formation, (4) the presence or complete absence of minimal desmoplasia, and (5) increased intratumoral lymphocyte infiltration (Fig. 1B). Finally, cases showing infiltration of scattered lymphocytes with prominent desmoplasia in the absence of lymphoid follicles, or with only one or two lymphoid aggregates per tissue section, were classified as CA (Fig. 1C).19 The prognosis was affected by the intensity and pattern of the inflammatory response. Among them, LELC cases had the best prognosis, followed by patients with CLR, who, in turn, had better survival rates than those with CA.17192930
Epstein-Barr Virus-Associated Immune Responses
Tumor microenvironment can play a critical role in patient outcome. TILs constitute the principal cellular component of the immune-active tumor microenvironment and their presence contributes to improved survival of patients with cancer. The number of TILs increases in EBVaGC, especially in LELC or CLR. Several studies found that EBVaGC is often accompanied by more extensive infiltration of CD8-positive cytotoxic T cells and a higher number of mature dendritic cells than EBVnGC.141530313233
Infiltrating immune cells at least partially contribute to antitumor immunity by promoting the eradication of EBV-positive malignant cells.193134 However, neither the mechanisms by which carcinoma cells are killed nor the biochemical cascades that allow them to evade the host immune response during tumor development and progression have been properly determined.
1. Cytokines
In 1998, transplantable human EBVaGC was designed and propagated in mice with severe combined immunodeficiency.35 The only cytokine gene that showed markedly higher expression in EBVaGC tumor strains compared to EBVnGC strains was IL1B that encodes interleukin-1β (IL-1β).36 IL-1β recruits numerous nonspecific lymphocytes to prevent direct contact between EBV-specific cytotoxic T cells and tumor cells.36
Using a cohort of GC RNA-Seq data sets from TCGA, Strong et al.37 performed a quantitative assessment of EBV gene expression in GC and analyzed EBV-associated alterations of cellular pathways. They found that EBV infection is associated with lower expression of tumor regulatory genes. Furthermore, samples with high levels of EBV gene expression displayed more extensive immune cell infiltration and higher interferon-gamma expression than samples with low or no EBV gene expression. These findings may explain the favorable prognosis of EBVaGC. On the other hand, high interferon-gamma levels induced the expression of indoleamine 2,3-dioxygenase (IDO1), a potent immune cell inhibitor, in EBVaGC cells. This circumstance may explain relative resistance of EBVaGC tumors to elevated numbers of immune cells.37
2. Deregulation of immune response genes
Kim et al.38 generated a gene expression profile analysis to compare tumor and non-tumor gastric tissues from 12 patients with EBVaGC and 14 with EBVnGC. It was revealed that EBVaGC cases had a higher degree of genetic homogeneity than EBVnGC cases. Deregulation of gene expression affected fewer genes in patients with EBVaGC than in those with EBVnGC. Furthermore, the majority of significant altered expression signals in EBVaGC are related to the immune response genes, especially the genes encoding proteins from cytokine (chemokine) pathways. These changes could recruit reactive immune cells to the intra- or peritumoral area of EBVaGC and this circumstance might contribute to longer survival of patients with EBVaGC compared to that of patients with EBVnGC.38
3. Epstein-Barr virus and programmed death receptor-ligand 1
At the genomic level, EBVaGC is accompanied by a relatively frequent recurrent amplification of the 9p24.1 locus containing the CD274 gene encoding PD-L1, which is also known as B7-H1.18 PD-L1 expression on tumor cells is known to play an important role in immune evasion. This is accomplished through the interaction with the co-inhibitory molecule programmed death receptor-1 (PD-1) expressed by T cells.39 The PD-1/PD-L1 interaction inhibits T lymphocyte proliferation, survival, and effector functions such as cytotoxicity and cytokine release. In addition, it induces apoptosis of tumor-specific T cells, promotes the differentiation of CD4+ T cells into Foxp3+ regulatory T (Treg) cells, and increases the resistance of tumor cells to cytotoxic T lymphocyte attacks.40 This interaction mediates the suppression of T cell functions, and thus reduces T-cell receptor (TCR)-mediated proliferation and cytokine production.39 Actually, we observed markedly increased levels of CD4- and CD8-positive T cells and a moderate increase of CD20 positive B cells around EBV-positive tumor cells. However, we observed very small numbers of CD1a positive dendritic cells (Fig. 2).
Blockade of PD-L1/PD-1 interaction may improve the efficacy of adoptive cell therapies in certain malignancies. Additionally, a correlation between prognosis and the level of PD-L1 expression has been reported in several human malignancies.414243444546474849505152535455 Although some controversy remains, many studies have shown that PD-L1 overexpression is associated with worse prognosis in many types of cancer.56 Two published studies of GC used immunohistochemical analysis and showed that abnormally high levels of PD-L1 are associated with poor prognosis,4142 while a recent study with 243 patients with curatively resected GC showed better disease-free survival and overall survival in patients with PD-L1 expression.57 In TCGA data, PD-L1/2 expression was elevated in EBVaGCs, in which IL-12-mediated signaling signatures induced robust immune cell presence.18 When coupled with the evidence of PD-L1/2 overexpression, this finding adds rationale for testing immune checkpoint inhibitors in EBVaGC.18
The findings described above do not offer complete explanation for all specific clinicopathological features of EBVaGC. Future studies are necessary to clarify the precise mechanisms of the immune reaction to EBV infection, additional clinical aspects of EBVaGC, and the therapeutic strategy for patients with EBVaGC.
Conclusion
One of the most distinctive clinicopathological features of EBVaGC is a prominent cellular immune reaction, reflected in the higher frequency of LELC and CLR compared to the occurrence of these signs in EBVnGC. This histological hallmark supports the hypothesis that interactions between tumor cells and tumor microenvironment play a critical role in the progression of EBVaGC. The intensity and pattern of the inflammatory response are strongly associated with the prognosis of GC. Accordingly, establishing the identity of cellular immune pathways affecting EBV infection during GC will be helpful for selecting appropriate targets for therapeutic intervention against EBVaGC. EBVaGC can be detected by a relatively simple method using EBER in situ hybridization; thus, any advances in the immunotherapy of EBVaGC can be readily extended to many patients with GC worldwide. Further studies are necessary to clarify the mechanisms underlying the cellular immune response in EBVaGC.

 XML Download
XML Download