

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Pertussis is one of the respiratory diseases caused by bacterial infection and specifically severe in the infants. The high global incidences of pertussis were reported until 1980's (1,982,384 cases/year)(1) but they were greatly reduced by vaccination campaign, which started from 1940. However, local outbreaks of pertussis were reported between 1990-2000 in well-vaccinated countries like USA, Netherlands, Japan, and Australia(2-4). Although there were many efforts to find out the reason of these outbreaks, they were not yet found. Therefore, to prevent the disease from spreading in early stage, the diagnostic process plays an important role.
Usually, the diagnostic gold standard is to confirm the pathogen by culture. However, the growth rate of Bordetella pertussis, the pathogen of pertussis, is quite slow. For this reason, the PCR detection is a critical step to determine the result in early phase. Until now, various kinds of diagnostic primer sets were developed. From these primer sets, BP1/2 for IS481 element(5, 6), PTp1/ p2 for pertussis toxin promoter region(5, 7, 8) and P1/2/3 for porin gene upstream region(9) are common primer sets used in clinical application. Especially, because of its high sensitivity and specificity, BP primer set is most widely used primer. However, despite of high sensitivity of BP primer, the cross-reactivity of BP primer set to B. holmesii was recently reported(10-13).
B. holmesii is one of the species of Bordetella Genus and shows highest similarity to B. pertussis in 16S rDNA sequence analysis(14). There were few reports about the pathogenicity and infectivity of B. holmesii for human, this strain was isolated from the nasopharyngeal specimens of patients with pertussis-like symptoms(14, 15). Moreover, the specific primer for B. holmesii was not reported yet and BP primers are currently applied to B. holmesii detection(10, 12, 13). Therefore, in diagnostic PCR diagnosis step, discriminative analysis between B. pertussis and B. holmesii is required.
For this reason, we designed new diagnostic primer sets for 4 major species of Bordetella Genus based on 16S rDNA sequence and the efficiency of designed primer sets was estimated.
MATERIALS AND METHODS
1. Bacterial Strains
The reference bacterial strains used in this study were Bordetella pertussis (ATCC 9797), Bordetella holmesii (ATCC 51541), Bordetella parapertussis (ATCC 15237) and Bordetella bronchiseptica (ATCC 31124). In addition to these strains, the nasopharyngeal specimens collected from pertussis patients were also used as confirmation test.
2. Preparation of PCR templates
The genomic DNAs of 4 Bordetella species were purified from cultured cells and used as PCR templates. For this, all of strains were incubated on Regan-Lowe medium without cephalexin for 5-7 days at 37°. After incubation, the bacterial cells were collected by scrapping and genomic DNA was isolated using commercial kit according to the manufacturer's instruction (QIAamp DNA Mini Kit, QIAGEN).
For clinical specimens, collected nasopharyngeal aspirates (NPA) were directly used as templates for PCR reaction. Some portion (0.5-1 mL) of the specimens was heated for 5 min in boiling water. Then after centrifugation, 1-2 µL of supernatant was used as PCR template.
3. PCR primers
The 4 PCR primers were designed from 16S rDNA sequences of 4 Bordetella species (B. holmesii, B. bronchiseptica, B. pertussis and B. parapertussis). The DNA sequence information of 16S rDNA gene was retrieved from NCBI GenBank. These collected nucleotide sequences were aligned using multiple alignment method by MEGA program(16) and the sequence similarity of 16S rDNA gene was calculated by p-distance method at Family level and Genus level(17). From these aligned data matrix, the region containing different nucleotide was analyzed by Sequence Output program and the PCR primers were designed by these differences (Fig. 1). For reference, the BP primers sequence (BP1:5'-GAT TCA ATA GGT TGT ATG CAT GGT T-3', BP2:5'-TTC AGG CAC ACA AAC TTG ATG GGC G-3') was cited from the published paper(12).
4. PCR condition
The standard reaction mixture (20 µL) was composed of 2 µL of 10x buffer, 0.5 µL of dNTPs mix (2.5 mM each), 1 µL of F-primer (5 pmoL/µL), 1 µL of R-primer (5 pmoL), 0.2 µL of SP-Taq polymerase (2.5 Unit/µL), 1 µL of template (purified genomic DNA or bacterial suspension) and distilled water was added to make total 20 µL of reaction volume. For 16S primer sets, dimethylsulfoxide (DMSO) was added to make 2.5% of final concentration.
The temperature condition for new 16S primer sets was 35 cycles of denaturation at 98° for 10 sec and annealing/extension at 64° or 66° for 15 sec according to primer sets. In the case of BP primer sets, 35 cycles of denaturation at 95° for 5 sec and annealing/extension at 54° for 10 sec were performed.
RESULTS
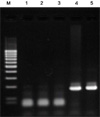
1. Cross reactivity of BP primer set with B. holmesii
The cross reactivity of BP primer to B. holmesii and other 2 Bordetella species (B. bronchiseptica and B. parapertussis) was confirmed in this study. As shown in Fig. 2, positive bands were appeared in both lanes (lane 4 and 5) of B. pertussis and B. holmesii. However, B. parapertussis (lane 2) and B. bronchiseptica (lane 3) were not detected by BP primers set. Therefore, the use of BP primers set in PCR diagnosis of pertussis should be considered.
2. Design of PCR primers sets
In this study, we considered the 16S rDNA gene to develop new primer set for PCR-diagnosis of pertussis. For this, the 16S rDNA sequences of 4 major Bordetella species (B. pertussis, B. holmesii, B. bronchiseptica and B. parapertussis) were collected from NCBI GenBank and aligned by multiple alignment method to find sequence variations. As the result, there were no specific variable regions. This indicates that 16S rDNA sequence was highly conserved in Bordetella species (98.7% of sequence similarity). However, the two positions of 661 bp and 782 bp in alignment matrix showed specificity according to their species. We selected these two sites as the motif for PCR primer design (Fig. 1) and 4 new primers were designed. As shown in Fig. 1, the designed primers were different in only one nucleotide of their 3'-termini.
3. Specificity of designed 16S PCR primer sets
As mentioned above, the specificity of designed new PCR primer sets was examined to 4 Bordetella species. For specific amplification, the combinations of these 4 primers were applied. 16S-F1/16S-R1 set for B. holmesii, 16S-F2/16S-R1 set for B. pertussis, and 16S-F2/ 16S-R2 set for other Bordetella species (Fig. 1) were used. As the result, the specific amplification of B. pertussis, B. parapertussis and B. bronchiseptica using the combinatorial primers sets was successfully performed. As shown in Fig. 3, the positive PCR bands appeared in only each target species (Lane 2, Lane 5, Lane 11 and Lane 12). Therefore, it was confirmed that the designed PCR primer sets showed the specificity to each Bordetella species.
16S-F2 and 16S-R1 primers were applied for B. pertussis. As shown in Fig. 3, the positive band was detected only in B. pertussis sample (Lane 5). However, we confirmed that the non-specific band was also detectable in the sample of B. holmesii under mild PCR condition. Therefore, stringent PCR condition was required in the use of 16S-F2 and 16S-R1 primers. For this reason, we added dimethylsulfoxide (DMSO) to reaction mixture until 2.5% of final concentration. As the result, 16S-F2/16S-R1 primers set specifically detected B. pertussis in this stringent condition.
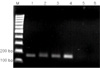
4. Sensitivity of designed 16S PCR primer sets
The sensitivity of these 2 primer sets (BP and 16S) for B. pertussis was estimated and compared. Under fixed PCR condition, the detection limit of target band was examined using purified genomic DNA. As shown in Fig. 4, when genomic DNA was used, the detection limit was 5 pg/reaction ((A)-lane 2). This estimated value was lower than the result of BP primer set (Fig. 4-(B)). BP gene was detectable as low as 10 fg of genomic DNA ((B)-lane 6).
In the case of cell suspension, as shown in Fig. 5, the detection limit of 16S rDNA gene was > 105 cells/ mL ((A)-lane 5). This result is not quite different with the result of BP gene. The detectable PCR band for BP gene was also shown from the cell suspension of > 104 cells/ml ((B)-lane 7). Therefore, these results indicated that 16S rDNA gene and BP gene showed similar detection range under practical diagnostic condition.
5. Application to clinical specimens
Applicability of new 16S primers to clinical specimens were estimated. The specimens were the nasopharyngeal aspirates collected from the suspected patients pertussis in 2006. These specimens were already confirmed positive for B. pertussis by BP primer PCR and culture. As shown in Fig. 6, all positive specimens were also confirmed positive by 16S PCR primer (Lane 1-4 in Fig. 6). Therefore, the newly developed 16S PCR primer sets were also applicable to the clinical specimens.
DISCUSSION
According to the WHO guide for Bordetella species diagnosis(1), the cross reactivity of BP primer to B. holmesii was already described. However, BP primer set is still used in most diagnostic laboratories because of its high sensitivity and specificity. Moreover, uncertainty of B. holmesii for human infection and carriage was also the reason. However, pathogenicity and carriage activity of B. holmesii for human were also proved recently(14, 15). For these reasons, there is need to develop new primer sets for discrimination of these two Bordetella species at PCR diagnostic step.
For this, we considered the 16S rDNA gene. As already known, 16S rDNA gene is most frequently used taxonomic marker in all Bacteria and Archaea(18). Firstly, we confirmed the sequence similarity of 16S rDNA gene to estimate their usefulness for B. pertussis. The group mean similarity of Bordetella Genus was 98.7% and it was higher than overall mean similarity of Alcaligenaceae Family (93.6%) containing Bordetella Genus. This indicates that 16S rDNA sequence is highly conserved to each Genus of Alcaligenaceae Family and this gene is a useful taxonomic marker to discriminate each Genus of Alcaligenaceae Family.
Currently, 9 Bordetella species have been reported in NCBI Taxonomy database (B. ansorpii, B. avium, B. bronchiseptica, B. hinzii, B. holmesii, B. parapertussis, B. petrii, B. trematum, B. pertussis). Among these species, the detectable species from human respiratory specimens were B. bronchiseptica, B. hinzii, B. holmesii, B. parapertussis and B. pertussis(19-21). Because B. hinzii was reported as rarely isolated species from immunocompromised(20) and cystic fibrosis patients(22), we did not considered it as major Bordetella species isolated from normal respiratory specimens. However, discrimination of B. pertussis and B. hinzii would be also possible by using new 16S primer sets (Fig. 1).
Although B. ansorpii and B. trematum were also isolated from human originated specimens(23, 24), they did not relate with human respiratory infection. Therefore, 4 Bordetella species (B. ansorpii, B. petrii, B. trematum, B. avium) were excluded from diagnostic targets in this study.
Especially, we also confirmed the specific primer set (16S-F1 and 16S-R1) for B. holmesii in this study (Fig. 3, Lane (2)). Until now, biochemical tests and cellular fatty acids analysis were mainly performed as identification tests for B. holmesii(14). In the molecular methods, DNA hybridization technique and 16S rDNA sequence analysis were performed(21, 25). However, there is no method for rapid identification of B. holmesii. IS481 based BP primer set for B. pertussis is used in B. holmesii detection(10, 12, 13). Therefore, there is need to develop PCR primer of B. holmesii for rapid identification. Although recA gene was recently reported as target gene for specific PCR of B. holmesii(26), other specific primer or target gene were not reported. As the result, the confirmed PCR primer set for B. holmesii in this report will be applicable to clinical diagnostic tests.
As shown in Fig. 4, 16S primer set showed 100 times lower sensitivity than BP primer set. It is attributed in the copy numbers of these two related genes. The BP primer was originated in IS481 repetitive sequence. It was known that there were approximately 80-100 copies of IS481 region in a bacterial cell(12, 27). However, for the 16S rDNA gene in the case of Tohama I strain, 3 copies per genome were present(28). Therefore, the major reason of reduced sensitivity of 16S primer was inferred as this different copy number.
As the conclusion, the specific PCR primers sets for B. pertussis, B. holmesii, B. parapertussis and B. bronchiseptica were developed based on 16S rDNA gene and their specific amplification of target strain was successfully confirmed in this study. Although the sensitivity was relatively low as compared to BP primers set, these new primer sets are applicable to clinical specimens and especially useful to discriminate B. holmesii and B. pertussis.




 XML Download
XML Download