

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Enterococci bacteria are facultative anaerobic grampositive cocci, which are considered part of the normal flora in humans and animals. However, these microorganisms may be the cause of several serious systematic infections too.123
The two most common Enterococcus species, E. faecalis and E. faecium are responsible for 80-90% and 5-10% of human enterococcal infections, respectively.2 E. faecalis is the most common isolate of nosocomial infections, but newly, due to increasing resistance to some antimicrobial agents, especially vancomycin, E. faecium isolates are also being considered.4 It has been shown that separate lineages of E. faecalis and E. faecium are leading causes of the large number of the multidrug-resistant enterococcal infections. According to several investigations, CC17 and closely related strains are the main agents of most hospital-acquired infections association with E. faecium.5 Although these organisms lack strong virulence factors, they may have an innate resistance/tolerance to many important antibacterial agents such as, cephalosporins, cotrimoxazole, and low levels of penicillin and aminoglycosides, polymyxin, lincosamide, trimethroprim-sulfamethoxazole, monobactams, streptogramin. They are also able to acquire resistance to penicillin, chloramphenicol, tetracyclines, aminoglycosides and vancomycin.3678
Pathogenicity and increased risk of acquisition of enterococcal infections is linked to antimicrobial resistance and expression of virulence factors including, adhesion factors, translocation, and immune evasion. In hospital settings, infective potential of enterococci can be due to selective the advantages conferred by their antibiotic resistance. Furthermore, there is great concern about infectious diseases because of spreading antimicrobial resistance genes. The most important infective agents of enterococci which have been identified, include: aggregation substances (asal), cytolysin (cyl), hyaluronidase (hyl), the enterococcal surface protein (esp) and gelatinase (gelE).910 It has been found that some of virulence factors such as, agg, esp and cyl genes, located on 153-kb pathogenicity island.11 Persistence of enterococci in the hospital setting may be associated with their virulence factors.11 Based on several studies, the virulence factors of gelatinase, aggregation substance and cytolysin have not been found in E. faecium in contrast to E. faecalis. On the other hand, esp and hyl have been found in both E. faecalis and E. faecium.12
Furthermore, clinical isolates of enterococci have virulence determinants that may result in promoting the emergence of infections and the persistence of these organisms in hospital settings which consequently can lead to increased resistance.13 This study was designed to scrutinize some virulence genes including asa1 (aggregation substance), esp (enterococcal surface protein), gelE (gelatinase), hyl (hyaluronidase) in clinical isolates of E. faecalis and E. faecium and to investigate possible correlations between virulence and antibiotic resistance.
MATERIALS AND METHODS
1. Identification of enterococcal isolates
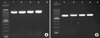
One hundred and seventy-five E. faecalis and sixty-seven E. faecium strains were collected from discrete clinical samples submitted to three teaching hospitals (including Beheshti, Besat and Farshchian Hospitals) located in Hamedan, Iran, from December 2012 to May 2014. The origins of the isolates were as follows: urine 200 (82.6%), endotracheal aspirate 23 (9.5%), blood 8 (3.3%), Skin soft tissue 6 (2.5%), and body fluids 5 (2.1%). The isolates were pinpointed using routine microbiological methods.14 Then, PCR targeting D-alanine- D-alanine ligases for E. faecalis (ddl E. faecalis) and E. faecium (ddl E. faecium) was used to confirm phenotypic speciation.15
2. Detection of E. faecalis and E. faecium species by PCR assay
Firstly, enterococcal DNA was extracted by boiling.16 Then, a mastermix PCR Kit [(PCR 2X Taq premix Mastermix), Ariatous Biotec Co.] was used to perform the PCR reaction. PCRs were performed with specific primers for each gene (Table 1) with some modifications to Kariyama's protocol15 using Eppendorf and Biorad thermocycler in a final volume of 20 µL. The thermal cycle program was performed by initial denaturation at 95℃ for 5 min, followed by amplification in 30 cycles of denaturation at 95℃ for 30 s, annealing at 52.5℃ for 30 s and elongation at 72℃ for 1 min, and a final extension at 72℃ for 10 min. E. faecalis ATCC 29212 and E. faecium BM4147 were used as quality control strains.
3. Antibiotic susceptibility testing
The antimicrobial susceptibilities of 175 E. faecalis and 67 E. faecium strains were examined by using the disk agar diffusion (DAD) method in accordance with the Clinical and Laboratory Standards Institute (CLSI) guidelines.1718 Erythromycin (15 µg), Tetracycline (30 µg), Ciprofloxacin (5 µg), Vancomycin (30 µg), Teicoplanin (30 µg), Norfloxacin (10 µg), Nitrofurantoin (300 µg), Quinopristin-Dalfopristin [Synercid (15 µg)] (Mast Co., UK), Chloramphenicol (30 µg), Gentamicin (10 µg), Linezolid (30 µg), and Ampicillin (10 µg) (HiMedia Mumbai Co., India) were used for antimicrobial susceptibility testing (AST).
In addition, minimum inhibitory concentrations (MIC) of the glycopeptide antibiotics i.e. vancomycin and teicoplanin (Sigma-Aldrich, Poole, Co., UK) against the E. faecalis and E. faecium isolates were determined using the microdilution broth method.1718 E. faecalis ATCC 29212 (Vancomycin sensitive), E. faecalis ATCC 51299 (vanB positive), E. faecalis E206 (vanA positive) were used as quality control.
4. Detection of infective genes esp, hyl, and asa1 by PCR
Multiplex PCR and single PCR were used for the identification of esp, asa1 and hyl virulence determinants using specific primers for each gene with some modifications on Vankerckhoven's protocol (Table 1).1319 Briefly, the final volume multiplex PCR reaction for genes esp and asa1, was 25 µL and for hyl gene was 20 µL. The PCR reactions were done for both mixtures on a Eppendorf and Biorad thermocycler (ASTEC Co., Japan) with an initial denaturation at 95℃ for 10 min, 30 cycles of amplification (denaturation at 94℃ for 1 min, annealing at 56℃ for 1 min, and extension at 72℃ for 1 min), and a final extension at 72℃ for 10 min.13 The E. faecalis ATCC 29212 (asa1 positive), E. faecium C68 (hyl and esp positive) were used as quality control.
5. Statistical analysis
Correlation between antibiotic susceptibility patterns and occurring virulence genes was analyzed statistically using the Chi-Square test; the differences were considered significant for p<0.0012 using the Bonferroni correction based on several primary comparisons. In addition, significant differences for simultaneous occurrance of virulence genes in enterococci strains was determined using Fisher's Exact test with a significance level of p<0.05. All tests were performed using SPSS software (version 19).
RESULTS
1. Isolation of enterococci strains
Using biochemical methods, of the 280 enterococcal isolates, 190 (67.8%), 75 (26.8%) and 15 (5.4%) isolates were recognized as E. faecalis, E. faecium and Enterococcus spp., respectively. Among the determined presumptive E. faecalis, E. faecium isolates, 175 (62.5%) E. faecalis and 67 (24%) E. faecium strains were confirmed using the PCR method (Fig. 1). Therefore, a total of 38 strains (13.5%) remained as a part of the Enterococcus genus and were excluded from the current study. Urine samples were the highest isolation source of E. faecalis and E. faecium strains. Among 175 E. faecalis strains, 153 isolates and among 67 E. faecium strains, 48 isolates were isolated from urine samples; followed by tracheal sample with 17 isolates (11 strains were determined as E. faecalis and 6 strains as E. faecium). The highest prevalence of E. faecium strains was observed in wounds and other organs samples (abscess and pulmonary secretions); among 12 strains isolated from wounds and other organs, 8 strains belonged to E. faecium (Table 2).
2. Antimicrobial susceptibility testing
Table 3 shows the susceptibility patterns of 67 E. faecium and 175 E. faecalis strains to 12 commonly used antibiotics. Resistance to the majority of antibiotics except for chloramphenicol, tetracycline, and quinopristin-dalfopristin was higher in E. faecium isolates compared to E. faecalis isolates. However, they both showed good rates of sensitivity to linezolid (100%), nitrofurantoin and chloramphenicol (74.6%). In fact, no resistance to the linezolid antibiotic was observed among E. faecalis and E. faecium strains; and all isolates of E. faecalis were susceptible to nitrofurantoin. Among 175 E. faecalis strains, the greatest resistance was observed for Synercid antibiotic with frequency of 167 (95.4%) isolates; followed by tetracycline with 154 (88%), erythromycin with 109 (62.3%), ciprofloxacin with 69 (39.4%), gentamicin with 63 (36%), Norfloxacin with 58 (33%), chloramphenicol with 57 (32.6%), vancomycin and teicoplanin with 9 (5%) and ampicillin with 6 (3.4%) isolates. Among 67 E. faecium isolates, the greatest resistance was observed for gentamicin with a frequency of 62 (92.5%) strains, followed by erythromycin with 58 (86.6%), norfloxacin with 56 (83%), ciprofloxacin with 54 (80.6%), vancomycin with 51 (76%), tetracycline and teicoplanin with 49 (73%), Synercid with 47 (70%), ampicillin with 42 (62.7%), nitrofurantoin with 17 (25.4%), and chloramphenicol with 7 (10.4%).
Of 175 E. faecalis isolates, resistances of 9 (5%) and sensitivity of 166 (95%) isolates to vancomycin and teicoplanin were confirmed by the microdilution broth method. Of 67 E. faecium isolates, 51 strains (76%) were resistant to vancomycin using the disk diffusion method, but resistance of 49 strains (73%) to vancomycin was confirmed by the Microdilution Broth method. 2 strains (3%) of E. faecium that were determined as resistant strains by disk diffusion using the Microdilution Broth method were identified as intermediate strains. In addition, resistance of 49 (73%) and sensitivity of 18 (27%) E. faecium isolates to teicoplanin using disk diffusion method were confirmed by the microdilution broth method. All 9 vancomycin-resistant E. faecalis (VREfs) and 49 vancomycin-resistant Enterococcus faecium (VREfm) strains had high-level resistances to vancomycin and teicoplanin, concurrently. The MIC value among 9 VREfs strains and VREfm strains was ≤512, 512.
3. Presence of infective genes
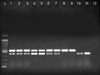
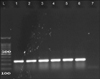
The distribution of infective factors among E. faecalis and E. faecium strains are presented in Table 4 and (Fig. 2 and 3). Among the E. faecalis strains, the asa1 gene was the most prevalent factor (97%), followed by the esp (78.3%) and hyl genes (56.6%); additionally, in E. faecium strains, the asa1 gene had the highest prevalence (100%) and hyl gene has the lowest frequency (71.6%), followed by the esp gene (82%).
In a total of 242 Enterococci strains, the asa1 gene was the most prevalent factor (98%), followed by the esp (79.3%) and hyl genes (60.7%). In the current study, the esp gene was found in 75% of the strains isolated from blood samples and 83% of strains isolated from wound infections.
Using statistical analysis, there was no significant difference for simultaneous occurrence of virulence genes in studied enterococci strains (p>0.05).
DISCUSSION
In the present study, we assessed antimicrobial resistance and three infective factors in clinically isolated E. faecalis and E. faecium and further analysis was conducted to scrutinize the relationship between the presence of virulence factors and antimicrobial resistances. In this study, there was a slight discrepancy between biochemical and PCR results for species identification, which is in accordance with other studies.2021 The main reason for this discrepancy is the similarities between Enterococcus species and the high phenotypic variation within individual species. PCR is a more accurate technique in comparison to the biochemical approach, so PCR results were preferred for the strains with discrepant identification.22 In this study, the esp gene was detected in 78.3% of E. faecalis and 82% of E. faecium isolates, this finding is similar to the results of other studies, which identified the esp gene in 47.1%,23 73%,24 68.4%7 of E. faecalis, and 80%,2526 65%,27 66%,23 71%,24 75%12 of E. faecium strains. However, this is in contrast to the findings of Shankar et al.28 and Channaiah et al.29 that reported the absence of esp in E. faecium. Although, as illustrated by Shankar et al.28 the esp gene was detected only in E. faecalis strains and other available studies have demonstrated a higher prevalence of the esp gene in E. faecalis,16 however, a study on the food and medical isolates indicated an increasing incidence of esp in clinical E. faecium isolates compared to E. faecalis.25
Willems demonstrated that the esp gene is a marker of the high prevalence of Enterococcus strains resistant to vancomycin in hospitalized patients,30 but according to the study of Woodford et al.,31 Sauer et al.,32 and Jahangiri et al.12 the esp gene also was identified in sensitive-vancomycin strains. In some studies, the prevalence of gene esp is reversed among VREfm and VSEfm strains. Whereas in other studies, the prevalence of gene esp was identical in VREfm and VSEfm strains. In accordance with the majority of these investigations, in the present study, the gene esp also was identified in a large number of VREfm strains (61%) compared with in VSEfm (21%). Camargo et al.33 demonstrated that esp was restricted to VREfm (56%) and not found in VSEfm. Vankerckhoven et al.10 surveyed virulence genes in eight European hospitals and found higher prevalences of esp in the clinical VREfm isolates (77%). Worth et al.34 and Sharifi et al.13 also found a higher incidence of esp in 80.5% and 71.05% in the clinical VREfm isolates, respectively. In the study by Terkuran et al.35 the esp gene was detected in 15.7% of VREfm isolates and the hyl gene in 28.6% of VREfm strains, but these genes were identified in 2.9% of Enterococcus sensitive and intermediate strains. In the study by Sauer et al.32 the esp gene was identified in 62.9% and 46.3% of VRE and VSE strains, respectively. Also, in the study by Jahangiri et al.12 esp gene was identified in 82% of clinical VREfm isolates and 53% of VSEfm isolates. The esp and hyl genes have significantly a higher prevalence among ampicillin-resistant VREfm isolates (53.7%, 37.3%) than ampicillin-susceptible VREfm isolates (19.4%, 22.4%); which is similar to the results of other studies.3334353637 The high frequency of esp gene, which was shown in the present study and most of the analogous studies, could be due to the fact that strains containing this gene can obtain antibiotic resistant genes and antibiotic resistant bacteria have long term stability in the body.2327
In the present study, the asa1 gene, (which encodes aggregation substance), was found in high frequency among E. faecalis (97%) and E. faecium strains (100%). A high incidence of this gene in E. faecalis was reported in previous studies. Results of studies on clinical E. faecium isolates are contradictory. In some studies, asa1 was not found in E. faecium but in contrast, in some studies this gene was detected in lower frequency and in our study and some other studies this gene was identified in higher prevalence among E. faecium isolates (Comerlato et al.38 and Kowalska-Krochmal et al.39), were detected it among 5%, 65% of VREfm and 2.7%, 60% of VREfs strains, respectively. Jahangiri et al.12 were not found asa1 gene in either 49 of VREfm strains or 17 of VSEfm strains. Sharifi et al.13 were detected asa1 gene in 80% of VREfs and 7.89% of VREfm strains. Hällgren et al.24 also were reported prevalence of asa1 in 79% of E. faecium strains. In studies by Huyckl & Gilmore, the asa1 gene was detected in 100% of blood isolates and 32% of non-blood isolates of Enterococcus. In studies by Elsner et al, Eaton & Gasson, and Archimbaud et al, the asa1 gene was detected in 40%-78% of clinical isolates of Enterococci.24 Hyaluronidase, coded by the chromosomal gene hyl, that influence the hyaluronic acid (hyaluronate, HA).35 Hyaluronidase in Enterococcus, indicates some homology to the hyaluronidases in other bacteria such as, Streptococcus pyogenes, Staphylococcus aureus and Streptococcus pneumoniae.9 We found the hyl gene among 49.3% of VREfm and 22.4% of VSEfm isolates, which is in accordance to findings of Rice et al.40 who detected the hyl gene among 71% of the United Kingdom VREfm isolates. But it was in contrast to the study by Jahangiri et al.12 which detected hyl gene among 80% of VSEfm and in 28.5% of VREfm isolates.
The reasons for the diversity in frequencies of hyl and asa1 can be as follows; Entrococcus strains are genetically different from each other based on their geographical origins. Moreover, the media which were used as sampling sources were varied between studies. In other words, some studies took their samples from blood, while others used urine, foods, or sewage.38263541 As demonstrated by Billström et al.19 and Wardal et al.42 the esp and hyl genes are linked with ampicillin and ciprofloxacin-resistant enterococci, particularly in CC17 which is an especially virulent, hospital adapted clone found globally. In the present study, it was shown that the esp virulence gene has a significant association with ciprofloxacin (p=0.001), erythromycin (p=0.001) and tetracycline (p=0.001) susceptibility patterns in E. faecium and with chloramphenicol (p=0.001) in E. faecalis strains; the hyl with teicoplanin (p=0.001) and vancomycin (p=0.001) in E. faecium strains; and also asa1 with vancomycin (p=0.001) in E. faecium and with ampicillin (p= 0.001) and chloramphenicol (p=0.001) in E. faecalis strains (Table 5).
The correlation between infective genes and antibiotic resistance in Enterococcus may vary from country to country. Based on the study by Hanna Billström et al.19 a significant relationship between imipenem, ampicillin, and ciprofloxacin resistance pattern and the espfm gene was found. Resistance to ciprofloxacin, imipenem, ampicillin antibiotics and the prevalence of the esp and hyl genes in these isolates were reported 90%, 80%, 77%, 56% and 4%, respectively. In the Study by Baylan et al.43 (Turkey, 2008) on E. faecalis strains, a significant association between the carriage asa1 gene and ciprofloxacin, norfloxacin and levofloxacin resistance pattern and between the esp gene and doxycycline resistance pattern were observed; in addition, a significant association between the hyl gene and nitrofurantoin resistance pattern in E. faecium strains was indicated. In the other study by Jankoska et al.41 there was no significant relationship between virulence genes and antibiotic resistance patterns; the esp gene was detected in 76% of isolates and all strains were susceptible to vancomycin and nitrofurantoin, 24%, 34%, and 28% of strains were resistant to ampicillin, ciprofloxacin and ceftriaxone, respectively.
According to the results of studies by Jahangiri et al.12 and Duprè,44 it was found that most of esp-positive isolates were resistant to more than 3 antibiotics. Lund et al.45 demonstrated that the existence of a strong correlation between the carriage of esp gene and antimicrobial resistance could be due to the higher conjugation frequencies in strains carrying the esp gene compared with strains lacking this gene. Sharifi et al.13 showed that E. faecium strains carrying the esp gene were resistant to more than 90% of the tested antibiotics and 64% of them were resistant to vancomycin. Considering these results, it seems that the esp gene facilitates E. faecium isolates ability to acquire antibiotic resistance genes. Van Wamel et al.46 showed that the expression level of the esp gene vary constantly between E. faecium strains depending on growth conditions and it is associated with bothe the initial connection and biofilm formation. Due to increasing resistance rates of enterococci to most common antibiotics, strict infection control measures are required. Antibiotic susceptibility testing is recommended for all patients before treatment for rational antibiotic use. There is a significant relationship between virulence genes and antibiotic resistance patterns. The virulence factors involve conjugative transfer of antibiotic resistance genes among enterococci strains and other species especially as the transfer of vancomycin resistance to staphylococcus aureus strains may occur.





 XML Download
XML Download