

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Bacteria-mediated cancer therapy (BCT) was first introduced in the late 19th century when William B. Coley used Streptococcus pyogenes to treat inoperable sarcomas and observed tumor shrinkage and increased patient survival.1 This is considered the first example of immunotherapy. During the next 40 years, Coley's toxins were given to over 1,000 cancer patients with excellent results, particularly in those with bone and soft-tissue sarcoma. However, his results were strongly criticized by the medical community, since his results were rarely repeatable. After the advent of radiation therapy and chemotherapy, Coley's toxins gradually disappeared from medical practice. However, recent progress in the fields of immunology and biotechnology has revived the mechanism underlying the activity of Coley's toxin, meaning that bacteria have returned to the agenda of those undertaking cancer research.
In the past 20 years, bacterial cancer therapy has again become a hot topic, and various kinds of bacteria have been the subject of preclinical and clinical research. Solid tumors have universal features, including abnormal blood vessels and hypoxic and necrotic regions;23 these microenvironments are suited to colonization by, and facilitate proliferation of, obligatory or facultative anaerobic bacteria such as Streptococcus,4 Clostridium,5 Bifidobacterium,6 Salmonella,789 Escherichia coli,101112 and Listeria.13 These bacteria specifically accumulate and proliferate inside tumor tissues, which can contain more than 1,000 times the number of bacteria found in reticuloendothelial organs like the liver or spleen.14 Attenuated Salmonella typhimurium (S. typhimurium) has been widely studied in this context due to its ready availability and ease of genetic manipulation. Different engineering strategies have been used to reduce bacteria-related toxicity, such as inhibiting expression of virulent genes. VNP20009, a S. typhimurium mutant strain (msbB-, purI-) harboring modified lipid A, and a purine auxotrophic mutation cause significantly less septic shock than their non-engineered counterparts and have been safely administrated to patients with metastatic melanoma and renal carcinoma in Phase I clinical studies; indeed, some bacterial colonization was observed in tumor biopsies.151617 A leucine/arginine auxotrophic strain, A1-R, effectively suppressed tumor growth in mouse models of cancer.81819 Another avirulent S. typhimurium strain defective in guanosine 5'-diphosphate-3'-diphosphate (ppGpp) synthesis, termed ΔppGpp,20 shows a good safety profile and has been used to treat mice bearing different cancers. 2122
S. typhimurium can grow under both aerobic and anaerobic conditions and so can colonize both large and small tumors. To increase therapeutic efficacy, bacterial therapy strategies were developed in combination with radiotherapy23 and chemotherapy,2425 or designed such that the bacteria delivered anticancer molecules.212627
KEY FEATURES OF S. TYPHIMURIUM THAT MAKE IT SUITABLE FOR CANCER THERAPY
Since the mid-20th century, radiotherapy and chemotherapy have developed rapidly and are now the main therapeutic regimens for cancer. However, neither destroys all cancer cells and both are toxic to normal tissues. The reasons for these side effects are as follows: (1) incomplete tumor targeting, (2) inadequate tissue penetration, and (3) limited toxicity to all cancer cells. These drawbacks limit treatment efficacy and are associated with increased morbidity and mortality.28 S. typhimurium have unique properties that can overcome these limitations: (1) the ability to sense and target tumors, (2) preferential growth in a tumor-specific microenvironment, (3) good intratumoral penetration, (4) low cytotoxicity and immunogenicity, and (5) versatile programmability.29 Salmonellae can be engineered as part of an active therapeutic approach to cancer and have multiple advantages over conventional therapies.
1. Sensing the tumor microenvironment
Hypoxia (<1% oxygen) is a near-universal feature of cancer, which makes it particularly resistant to radiotherapy and chemotherapy.3 Due to rapid cell proliferation and continuous angiogenesis during tumor growth, the newly formed blood vessels are premature both in terms of the endothelial cell lining and binding ends, resulting in sluggish blood flow and insufficient delivery of oxygen and nutrients to the tumor tissue. Oxygen is one of the most important signals used by bacteria to sense the tumor microenvironment. Low oxygen concentrations favor tumor-specific colonization and proliferation of facultative anaerobes such as S. typhimurium. The characteristic of hypoxia has been fully utilized to increase tumor-specific accumulation of bacteria expressing the essential asd (aspartate-semialdehyde dehydrogenase) gene under hypoxic conditions,30 or via delivery of therapeutic drugs under the control of hypoxia-inducible promoter-1 (HIP-1),31 thereby reducing cytotoxicity to normal tissues and increasing target specificity. Bacterial chemotaxis in response to small nutrient molecules such as ribose and amino acids, which are released from dying tumor tissues, occurs in tumors both in vitro and in vivo.3233 Genetically engineered auxotrophic strains of S. typhimurium (such as purine, leucine, and arginine auxotrophs) show increased tumor-specific accumulation.141934
2. Tumor penetration and proliferation
Attenuated bacteria are metabolically active and able to perform specific activities within tumor tissues, which make them different from chemical and biological drugs. S. typhimurium have properties that enable them to better penetrate tissue than chemical drugs. Motility is a key feature of bacteria that enables them to actively swim away from chaotic vasculature and spread throughout tumor tissues.35 The motility of attenuated Salmonellae has also been harnessed to enable delivery of encapsulated microbeads to tumors with theranostic molecules.36 In additional to motility, bacterial metabolism and host macrophages also play important roles in the bacterial distribution and tumor colonization.37 Depletion of macrophages resulted in enhanced tumor colonization and increased bacterial upload in spleens, while aromatic amino acid biosynthesis-deficient strains showed a higher specificity to tumors. S. typhimurium specifically accumulate and proliferate in tumor tissues, resulting in bacterial numbers that are over 1,000-fold higher (as high as 1010 cfu/g tissue) than those in normal tissues such as liver and spleen tissue.143338 Bacterial colonization of tumor tissues deprives cancer cells of nutrients and activates antitumor immunity, leading to tumor cell death.39
3. Immune stimulation
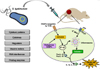
Tumors are immunosuppressive in nature and escape immune surveillance by limiting the maturation and infiltration of immune cells.3940 Systemic administration of attenuated S. typhimurium activates anticancer immunity, leading to tumor regression. Specifically conserved bacterial components, called pathogen-associated molecular patterns (PAMPs), are recognized by pattern recognition receptors (PRRs) known as toll-like receptors (TLRs), which broadly recognize molecules shared by pathogens, but not host cells/tissues. Bacterial components such as flagellin, LPS, or CpG sites are strong agonists for TLRs and activate TLR signaling pathways to induce innate and adaptive immune responses.738 We recently reported that systemic infection by attenuated S. typhimurium also leads to the activation of inflammasome pathways during the early stages of bacterial colonization of cancer tissues through the NOD-like receptor (NLR) family.38 IPAF inflammasomes recognize cytoplasmic bacterial flagellin injected via bacterial secretion systems,41 whereas NLRP3 inflammasomes are activated by endogenous danger signals (damage-associated molecular pattern molecules) or PAMPs along with increased K+ efflux.42 Activation of caspase-1 by inflammasomes results in the cleavage of pro-IL-1β and pro-IL-18 to yield active IL-1β and IL-18, respectively (Fig. 1).738 In summary, systemic administration of attenuated S. typhimurium induces the production of proinflammatory cytokines such as IL-1β, IL-18, TNF-α, and IFN-γ, resulting in recruitment and activation of immune cells, including dendritic cells, macrophages, and T cells, that convert the tumor microenvironment from immunosuppressive to immunocompetent.738
4. Programmability
Attenuated bacteria were have been used as a delivery vehicle to express cargo molecules, such as cytotoxic agents, 21223031434445 cytokines,464748495051 RNA interference,525354555657 and prodrug enzymes,17585960616263 in tumors to enhance BCT (Fig. 1). The expression of cargo molecules could be precisely regulated by external signals to reduce unwanted expression in normal tissues. To date, two categories of gene-triggering systems have been investigated in attenuated S. typhimurium-mediated cancer therapy: 1) external trigger systems such as the L-arabinose-inducible pBAD promoter,2164 tetracycline- or doxycycline-inducible pTet promoter22, and γ-irradiation-inducible pRecA promoter;23 and 2) environmental sensing systems including the hypoxia-inducible fumarate and nitrate reduction regulator,3165 and the quorum-sensing system, which turns on transgene expression at high bacterial densities that are usually observed in tumor tissues.66 We previously showed that early induction of therapeutic gene expression from the moment of bacterial injection may damage normal organs, including the liver and spleen (as identified by clinical chemistry parameters). We evaluated systemic toxicity by administration of doxycycline on the day of (0 dpi) and 3 days (3 dpi) after bacterial infection, when the tumor was colonized by S. typhimurium expressing cytolysin A (ClyA) under the control of the pTet promoter. The results showed that improper induction of ClyA (0 dpi) induced high levels of alanine aminotransferase and aspartate aminotransferase, indicating injury to reticuloendothelial organs. However, proper induction of ClyA (at 3 dpi) prevented hepato- and spleno-toxicity.22 To increase tumor-specific targeting capabilities, bacteria were genetically engineered to display tumor-targeting moieties on the cell surface. Attenuated S. typhimurium displaying a single-domain from an antibody against tumor-associated antigen (CD20) specifically infected CD20-positive lymphoma cells, with few nonspecific effects.67 The non-invasive S. typhimurium strain ΔppGpp, which is engineered to express the RGD peptide on its surface, selectively targeted and killed xenograft cancer cells overexpressing αvβ3 integrin, and is considered a novel and promising approach to the development of bacteria-mediated delivery systems.68
ATTENUATED STRAINS FOR CANCER THERAPY
Various S. typhimurium mutant strains have been studied from the perspective of cancer treatment; these are summarized in Table 1. Different strategies have been used to engineer bacteria to reduce cytotoxic effects in normal organs and increase specific colonization of tumors.
1. VNP20009
This is an attenuated S. typhimurium strain that has been widely studied in animal cancer models and tested in Phase I clinical studies involving human cancer patients. VNP20009 is a genetically modified S. typhimurium strain possessing an excellent safety profile, with a 10-fold reduction in TNF-α production compared to wild type. The msbB mutant strain showed greatly reduction of TNF-α-induced septic shock.33 To improve tumor-specific colonization, VNP20009 was further genetically engineered by depleting the purI gene. Because the growth of VNP20009 depends on the level of purine, they are more likely to colonize and proliferate in purine rich regions, such as tumor tissues. VNP20009 was examined in various murine cancer models, including melanoma,9 breast cancer,23 and colon cancer,46 and in a canine model of spontaneous neoplasia.69
2. A1-R
The A1-R strain was developed using nitrosoguanidine mutagenesis. This leucine and arginine auxotrophic strain grew more strongly in neoplastic tissues than in normal organs, and the colonizing bacteria could be re-isolated from tumor tissue.34 A1-R inhibited the growth of different cancers in mouse models of prostate cancer,18 breast cancer,19 cervical cancer,24 and glioma;70 it also inhibited metastasis. 871 The attenuated strain also "tricked" quiescent cancer cells into the cell cycle from G0/G1 to S/G2/M, thereby increasing the sensitivity to chemotherapy.72 Alteration of the cell cycle is a novel strategy for combinational cancer therapy.
3. ΔppGpp
Another avirulent S. typhimurium strain, ΔppGpp, was engineered by regulating endotoxin gene expression. A double mutant Salmonella strain (relA-, spoT-) defective in ppGpp synthesis, resulting in downregulation of endotoxin gene expression, is avirulent in mice after systemic infection, showing an approximately 105,6-fold increase in the LD50 values when compared with the wild type after oral administration or intraperitoneal inoculation.73 ΔppGpp achieved excellent tumor suppression by activating the inflammasome pathway, as observed by upregulated expression of genes encoding inflammasome-related NLRP3, IPAF, and tumor inhibitory cytokines (IL-1β and IL-18, TNF-α), thereby suppressing the proliferation of CT26 and MC38 colon cancer models in mice.738 The ΔppGpp strain is also a good vector for the tumor-specific delivery and expression of therapeutic molecules. The engineered bacteria can also express imaging reporter genes such as firefly luciferase and renilla luciferase,2274 or tumoricidal agents such as cytolysin2122 and the mitochondrial-targeting domain (MTD) of Noxa.26
4. Other strains
The three strains described above represent attenuated S. typhimurium, which has been well studied with respect to cancer. Three basic mechanisms are involved in the creation of mutant strains. First, modification of bacterial components to reduce the induction of inflammation, e.g., removal of lipid A from VNP20009 (msbB-, purI-);14334669 second, creation of nutrient auxotrophs by depleting certain genes to enable the bacteria to survive and proliferate in tumor tissues (e.g., leucine and arginine auxotrophic A1-R (Leu-, Arg-),1819707172 aromatic compound-dependent BRD509 (aroA-, aroD-),4775 adenine-dependent MvP728(purD-, htrA-),52 and strains derived from SL7207 (aroA-) that express the essential asd gene under hypoxic conditions3076); third, creation of strains by inactivating or downregulating expression of endotoxin-related genes (including ΔppGpp (relA-, spoT-),72173 LH340 (phoP-, phoQ-),5377 and SB824 (aroA-, sptP-)).78 To engineer such bacteria with better performance, we usually combine different strategies, for example, to yield the lipid A mutant VNP20009 that is defective in purine synthesis. Also, nutrient auxotrophs always show downregulated expression of endotoxin genes.
STRATEGIES FOR S. TYPHIMURIUM-MEDIATED CANCER THERAPY
Attenuated S. typhimurium suppress various cancers in mouse models. Different strategies have been developed to increase their effectiveness, including combinational therapy with radiation or chemical drugs and genetic engineering of bacteria to express therapeutics such as cytotoxic proteins, cytokines, prodrug enzymes, regulators, and genetic materials used for DNA vaccine or gene silencing (Table 2).
1. Native cytotoxicity and combinational therapy
Attenuated S. typhimurium effectively inhibits tumor growth.78934707172 Native bacterial cytotoxicity is mediated by activation of the host immune system and by depriving cancer cells of nutrients.39 Bacterial components (such as LPS, flagellin, and CpG) and signals/molecules released from damaged cancer cells activate the TLR and NLR signaling pathways, resulting in the production of proinflammatory cytokines (IL-1β, TNF-α, and IL-18), which mediate an antitumor immune response. In addition, rapidly proliferating bacteria deprive tumors of nutrients, resulting in cancer cell starvation and death. Administration of attenuated S. typhimurium, VNP20009, to 41 dogs with spontaneous neoplasia resulted in tumor colonization in 42% cases (as demonstrated by tissue culture). Moreover, 15% of dogs mounted a strong antitumor response (four showed a complete response and two a partial responses).69 Bacteria have been used in combination with other regimens to generate synergistic antitumor effects by increasing the sensitivity of cancer cells to other therapies and by reducing toxicity. Bevacizumab (Avastin, BEV) is a humanized monoclonal antibody that targets the vascular endothelial growth factor (VEGF) receptor, thereby inhibiting angiogenesis; this antibody is widely used to treat cancer patients, although many tumors acquire resistance after a short period of response.79 Combined treatment of nude mice harboring orthotopic human pancreatic tumors with BEV and an attenuated A1-R strain led to a reduction in tumor weight.80 The administration of a traditional Chinese herbal mixture along with A1-R enabled bacterial administration at large doses, which increased antitumor efficacy and reduced toxicity when compared with monotherapy with bacteria alone.25 We recently reported that therapy with engineered E. coli led to a significant improvement in the results achieved by radiotherapy, and that this combination produced a better treatment outcome with only 1/3–1/2 of the conventional radiation dose.10 While hypoxic cells within malignant tumors are often resistant to radiotherapy, facultative anaerobic bacteria proliferate in hypoxic areas and kill cancer cells. Thus, a combination of radiotherapy and bacteriotherapy may make up for the weaknesses of each therapeutic modality alone.
2. Cytotoxic proteins
Attenuated S. typhimurium can be used as a vector to deliver and express tumor-specific cytotoxic agents to retard tumor growth. However, expression of toxic genes must be tightly regulated by inducible or tumor-specific promoters to avoid unintended damage in normal tissues.21223031 Cytolysin A (ClyA, HlyE) is a native bacterial toxin produced by E. coli, Salmonella enterica serovar Typhi, and Paratyphi A, and is cytotoxic to cultured mammalian cells due to its pore-forming activity. A ΔppGpp strain engineered to produce ClyA effectively suppressesed the tumor growth in various cancer models.212230 Proapoptotic ligands such as Apoptin,43 Fas ligand, and TNF-related apoptosis-inducing ligand (TRAIL) selectively induced apoptosis via death receptor pathways, resulting in tumor suppression and prolonged survival.4445
3. Cytokines
Attenuated S. typhimurium have been engineered to deliver immunocompetent cytokines such as IL-2, IL-18, CCL21, and LIGHT for targeted cancer immunotherapy.464748495051 Tumor-specific cytokines produced by bacteria kill cancer cells by triggering the host immune system via upregulation of immune cell activation, proliferation, and migration. IL-2 is a signaling molecule that regulates lymphocyte activity. IL-2-induced tumor suppression correlates with reduced angiogenesis and increased necrosis within tumor tissues.47 IL-18 (also known as IFN-γ-inducing factor) increases the cytolytic activity of T cells and NK cells, along with cytokine production.49 Furthermore, IL-18 upregulates MHC class I antigen expression and drives the differentiation of CD4+ helper T cells into Th1 cells, and suppresses angiogenesis by inhibiting the proliferation of endothelial cells, thereby amplifying the antitumor effects mediated by NK cells, macrophages, and CD8+ T cells.49 CCL21 controls the migration of lymphocytes, dendritic cells, and NK cells.4650 LIGHT (also known as TNFSF14 or HVEM-L) is a TNF family cytokine that is homologous to lymphotoxin and binds both the lymphotoxin-β receptor (LTβR), which is expressed on epithelial cancers, and herpes virus entry mediator (HVEM), which is expressed by T lymphocytes. Attenuated S. typhimurium encoding LIGHT showed promising antitumor effects in immunocompetent mice bearing syngeneic tumors.51
4. Regulators
Certain cargo molecules were designed to regulate tumor cell growth or inhibit angiogenesis in tumor tissues. Sprouty (SPRY1/2) proteins are endogenous negative regulators of receptor tyrosine kinase (RTK) signaling pathways. Activation of RTK signaling pathways often correlates with cancer cell proliferation, angiogenesis, and progression. Delivery of SPYR to tumor tissues via engineered VNP20009 significantly inhibited melanoma growth in vivo; tumor suppression was mainly mediated via inhibition of ERK1/2 phosphorylation.81
p53 is crucial for apoptosis, genomic stability, and inhibition of angiogenesis and carcinogenesis. The activity of p53 is negatively regulated by mdm2 (murine double minute 2) and by ubiquitin-mediated degradation.82 Activation of p53 induces expression of downstream target genes related to cell cycle arrest and apoptosis. The E6 viral oncoprotein binds to wild-type p53 (wt-p53) in the host cell and disrupts its function; thus gene silencing of E6 in HPV-positive cervical cancer restores p53 function with respect to cell cycle arrest and apoptosis, leading to cancer suppression both in vitro and in vivo.82 Also, engineered Salmonella co-expressing p53 and mdm2 siRNA increased the therapeutic effect of cisplatin against prostate cancer.54 A Δ ppGpp strain expressing the MTD of Noxa induced cell death by increasing cytosolic calcium concentrations and mitochondrial permeability.26 Also, angiogenesis inhibitors such as endostatin and ENDO-VEGI115 inhibit tumor angiogenesis when expressed locally by attenuated Salmonella. 565777
5. Vaccine vectors
Attenuated S. typhimurium is widely used as a DNA vaccine vector. Bacteria-mediated delivery of cancer-specific antigens or antibodies, growth factor-targeting domains, and anti-apoptosis or tumor-associated macrophage-targeting proteins can stimulate the immune system, promote inflammation, and increase antigen presentation to T cells. Most DNA vaccines are orally administered and have the potential to inhibit or prevent tumor growth when given in multiple doses. Survivin, which is highly regulated and optimally expressed during the G2/M phase of the cell cycle, is essential for anti-apoptotic function. A DNA vaccine encoding survivin could induce CD8 T cell-mediated anticancer activity.508384 Endoglin (CD105) is a coreceptor for the TGF-β receptor complex and is overexpressed on proliferating endothelial cells; thus it is an attractive target for antiangiogenesis-mediated cancer therapy.2785 A 4-1BB ligand (4-1BBL) DNA-based vaccine effectively suppressed colorectal cancer development in rats by increasing T cell-mediated immunity.86 Oral vaccines encoding murine VEGF receptor-2 (flk1) inhibited angiogenesis and showed antitumor effects.8788 Cancer-specific antigens such as the carcinoembryonic antigen (CEA), the human papillomavirus antigen (HPV16 E7), and cancer antigen-specific antibody domains, such as including a CEA antibody-specific chain and a tumor-associated antigen CD20-targeting antibody, have been expressed and shown antitumor activity.67758990 A DNA vaccine against MTDH/AGE-1 (astrocyte elevated gene-1, which is overexpressed in >40% of breast cancer patients and is known as metadherin (MTDH) in mice) significantly suppressesed lung metastasis of breast cancer and increased chemosensitivity to doxorubicin treatment.91 Immunostimulating molecules such as glycoprotein (gp100) or heat shock protein (M. tuberculosis heat shock protein 70) elicit host-derived protective immunity to suppress tumor development.9293
6. RNA interference
Engineered bacteria have been used to transfer genetic material for the targeted silencing of certain genes to inhibit tumor growth and increase sensitivity to therapy. Gene-specific small hairpin RNAs (shRNAs) delivered by S. typhimurium are processed by the enzyme dicer to yield small interfering RNAs (siRNAs) that trigger degradation of the target RNA.53 The signal transducer and activator of transcription (Stat3), a factor that inhibits apoptosis and promotes cell growth and is overexpressed in many kinds of cancer, is an attractive target for shRNA-mediated gene silencing and has been widely studied with respect to the prevention of metastasis and inhibition of tumor growth.52535694 S. typhimurium transformed with an immunosuppressive molecule (indoleamine 2,3-dioxygenase; IDO)-targeting shRNA suppresses the growth of pancreatic cancer and melanoma in mouse models.9596 Targeting survivin or multidrug-resistant gene (MDR1) may significantly inhibit tumor growth and increase sensitivity to chemotherapy.555797 Silencing of mdm2 rescues the activity of p53 to regulate cell cycle and function, thereby suppressing tumor growth.54
7. Enzymes
Enzymatic proteins expressed by engineered S. typhimurium can convert nontoxic prodrugs into toxic anticancer drugs in cancer tissues, thereby minimizing systemic toxicity. Administration of attenuated S. typhimurium expressing E. coli purine nucleoside phosphorylase (ePNR) converts two prodrugs, 6-methylpurine 2'-deoxyriboside (MePdR) and 6-methoxypurine 2'-deoxyriboside (MoPdR), into toxic substances named 6-methylpurine (MeP) and 6-methoxypurine (MoP), leading to tumor-specific cell killing.585960 The prodrug activation enzyme, carboxypeptidase G2 (CPG2), activates different prodrugs and induces cytotoxicity in human tumor cells, but not in the host bacterium.61 Attenuated VNP20009 expressing E.coli cytosine deaminase (CD) have been injected directly into the tumors of cancer patients; this enzyme converts 5-fluorocytosine (5-FC), an antifungal agent with limited systemic toxicity, into 5-fluorouracil (5-FU), a cytotoxic anticancer drug commonly used in the clinic to treat head and neck, gastric, colorectal, pancreatic, and breast cancers. The tumor to plasma ratio of 5-FU was 3.0, whereas that of noncolonized tumors was less than 1.0.17 Bacteria harboring enzymes are also used for positron emission tomography (PET)-based reporter gene imaging. The VNP20009 encoding herpes simplex virus thymidine kinase (HSV1-tk) selectively colonized tumor xenografts and effectively sequestered a radiolabeled nucleoside analogue, 2'-fluoro-1-β-D-arabino-furanosyl-5-iodouracil (FIAU), thereby facilitating BCT and diagnostic imaging.6263
PREVIOUS CLINICAL STUDIES
Despite the fact that Coley's toxins were used to treat cancer patients over 100 years ago, the application of S. typhimurium for cancer treatment was first reported after the year 2000. Used in abundant preclinical studies in animal models ranging from mice to monkeys, VNP20009 was designed by Vion Pharmaceutics Inc. and tested in 25 human patients (24 with metastatic melanoma and one with metastatic renal carcinoma).15 The maximum-tolerated dose (MTD) was estimated as 3.0×108 cfu/m2. Dose-related toxicity was due to increased systemic production of proinflammatory cytokines, including IL-1β, TNF-α, IL-6, and IL-12. Focal bacterial colonization of tumors was observed in two patients receiving 1.0×109 cfu/m2 and in one patient receiving 3.0×108 cfu/m2. However, no objective tumor regression was observed, even in patients with colonized tumors. Further clinical studies involved four additional metastatic melanoma patients.16 VNP20009 was given at a MTD of 3.0×108 cfu/m2, resulting in minor and transient side effects during therapy. Only one colony was detected in an excisional tumor biopsy obtained within 2 weeks of therapy. One month after therapy, two patients showed significant disease progression and were thus treated with other therapeutic regimens. The third patient was free of disease after surgery and was still disease free at the 3 month follow-up appointment. While the fourth patient showed initial tumor suppression (both in terms of tumor size and the number of subcutaneous lesions) the disease progressed in the next 2 months.
To improve the therapeutic efficacy, another preclinical study used a modified strategy in that attenuated VNP20009 expressing E. coli CD were injected directly into tumors.17 CD converts 5-FC to toxic 5-FU. Three patients with squamous carcinoma and adenocarcinoma were treated by intratumoral injection of these bacteria (doses ranged from 3×106 to 3×107 cfu/m2 once every 28 days as long as no disease progression or intolerable toxicity was observed). No adverse reaction was noted after six cycles of treatment. Two patients showed bacterial colonization in tumors for at least 15 days after initial administration. The conversion of 5-FC into 5-FU (tumor to plasma 5-FU ratio) in the two colonized patients was 3.0, whereas that in the noncolonized patient was less than 1.0. The results suggest that attenuated S. typhimurium can deliver therapeutic genes to malignant tissues at safe doses.
CONCLUSION AND FUTURE PERSPECTIVES
Engineering of attenuated bacteria for targeted cancer therapy has been tested in animal models and promotes significant tumor suppression and prolonged survival. Facultative anaerobic BCT have some advantages over other therapies, including self-propulsion, self-proliferation, environmental sensing, external detection, and convenience in gene modification.29 These characteristics make bacteria an ideal and novel strategy for targeted cancer therapy. Various bacteria have been evaluated; among them, S. typhimurium is one of the most promising and the first-in-human studies to suggest the possibility of clinical translation.
Attenuated S. typhimurium reduced endotoxin expression and increased tumor-specific colonization, enabling administration of bacteria at relatively high doses to achieve tumor suppression. Systemic infection by attenuated bacteria results in a greater than 1,000-fold increase in the number of bacteria in the tumor tissue relative to that in normal organs such as the liver and spleen. Different strategies have been used to engineer bacteria to express tumor-inhibiting proteins,2174 transfer eukaryotic expression vectors to infected cancer cells,3058 and deliver targeted gene silencing.5257
Phase I clinical studies of VNP20009 in cancer patients have shown that tumor targeting is a major obstacle to further development. Future studies should focus on improving bacterial-targeting efficiency. Such studies may go in two possible directions: one is to use tumor-amplified protein expression therapy (TAPET™) to maximize the benefits derived from bacterial cancer therapy;1762 the other is bacterial surface engineering to display cancer-targeting domains. For example, the RGD peptide binds to αvβ3, which is overexpressed by various cancers, thereby increasing tumor-specific colonization.68
Optimal bacterial strains should be selected for different strategies. For example, non-invasive bacteria would better trigger cell surface receptors or PRRs, while invasive bacteria would be better for delivery of cancer-specific antigens to immune cells. The target cell is also of great importance because immune cell-targeting bacteria and cancer cell-targeting bacteria may have different roles. Moreover, the development of BCT requires multidisciplinary collaboration among those working in the fields of microbiology, immunology, cancer biology, chemical engineering, medical imaging, radiation oncology, and clinical oncology.


 XML Download
XML Download