

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Interstitial fibrosis and tubular atrophy of the kidney is the final common pathogenesis of chronic kidney disease. Unilateral ureteral obstruction (UUO) is a well-known model for progressive renal disease. Prolonged ureteral ligation induces marked hydronephrosis of the obstructed kidney, which exhibits tubulointerstitial fibrosis and apoptosis in association with activation of pro-fibrotic and pro-apoptotic proteins.1
The mammalian target of rapamycin (mTOR) is a serine/threonine kinase that plays diverse functions in development, metabolism, aging, cellular proliferation, and cell growth.2-4 Recent data have demonstrated that the mTOR signaling pathway is involved in the pathogenesis of kidney diseases such as acute kidney injury, diabetic nephropathy, and polycystic kidney disease.5-7 Activation of the mTOR pathway in diabetic nephropathy is associated with interstitial fibrosis, tubular atrophy, and progressive decline of the glomerular filtration rate, and blockade of the mTOR pathway slows the progression of diabetic nephropathy.8-10 In addition, we demonstrated that phosphorylation of the mTOR signaling pathway was increased in the kidney of deoxycorticosterone acetate (DOCA)-salt hypertensive rats.11 However, the role of the mTOR signaling pathway in the pathogenesis of kidney injury of the ureteral obstructed kidney has not been fully elucidated.
The present study aimed to investigate changes in the mTOR signaling pathway in the pathogenesis of tubulointerstitial fibrosis and apoptosis in the obstructed kidney of rats with UUO.
MATERIALS AND METHODS
1. Animals
The animal study was approved by the Ethics Committee of Chonnam National University Medical School. The experimental procedure conformed to the institutional guidelines for experimental animal care and use. Male Sprague-Dawley rats weighing 200 to 220 g were used.
2. Induction of UUO
Rats were unilaterally obstructed by ligation of the left proximal ureter for 7 days. Control rats were treated in the same way except that no ligature was made. The rats had free access to standard rat feed and tap water and were sacrificed by decapitation on day 7 after operation. The kidney was rapidly removed, dissected into three zones [cortex and outer stripe of outer medulla (cortex/OSOM), inner stripe of outer medulla (ISOM), and inner medulla (IM)], and processed for semiquantitative immunoblotting as described below.
3. Semiquantitative immunoblotting
The dissected cortex/OSOM, ISOM, and IM were homogenized in ice-cold isolation solution containing 0.3 M sucrose, 25 mM imidazole, 1 mM EDTA, 8.5 µM leupeptin, and 1 mM phenylmethylsulfonyl fluoride, with pH 7.2. The homogenates were centrifuged at 1,000×g for 15 min at 4℃ to remove whole cells, nuclei, and mitochondria. The total protein concentration was measured (Pierce BCA protein assay reagent kit, Rockford, IL, USA). All samples were adjusted with isolation solution to reach the same final protein concentrations and solubilized at 65℃ for 15 min in SDS-containing sample buffer and then stored at -20℃. To confirm equal loading of protein, an initial gel was stained with Coomassie blue. SDS-PAGE was performed on 9% or 12% polyacrylamide gels. The proteins were transferred by gel electrophoresis (Bio-Rad Mini Protean II, Bio-Rad, Hercules, CA, USA) onto nitrocellulose membranes (Hybond ECL RPN3032D; Amersham Pharmacia Biotech, Little Chalfont, UK). The blots were subsequently blocked with 5% milk in phosphate-buffered saline with Tween 20 (PBST; 80 mM Na2HPO4, 20 mM NaH2PO4, 100 mM NaCl, 0.1% Tween 20, pH 7.5) for 1 hour and incubated overnight at 4℃ with 1:1,000 dilutions of primary antibodies, followed by incubation with 1:1,500 dilutions of secondary anti-rabbit horseradish peroxidase-conjugated antibodies (P448; DAKO, Glostrup, Denmark). The labeling was visualized by use of an enhanced chemiluminescence system.
4. Primary antibodies
The antibodies used were as follows: affinity-purified anti-rabbit antibodies against phosphorylated phosphatidylinositol 3-kinase (p-PI3K; Cell Signaling Technology, Beverly, MA, USA), phosphorylated Akt (p-Akt; Cell Signlaing Technology), phosphorylated mTOR (p-mTOR; Cell Signlaing Technology), transforming growth factor-β1 (TGF-β1; Santa Cruz Biotechnology, Santa Cruz, CA, USA), Bax (Cell Signaling Technology), and Bcl-2 (Cell Signaling Technology).
RESULTS
1. Weight of body and kidney
On day 7 after the ureteral obstruction, the body weight of rats did not differ significantly between the two groups (238±5.8 g in controls vs. 240±4.5 g in UUO, p=0.79). However, the weight of the ureteral obstructed kidney was increased compared with the controls (1.2±0.1 g in controls vs. 2.3±0.2 g in UUO, p<0.05).
2. Expression of mTOR signaling pathway
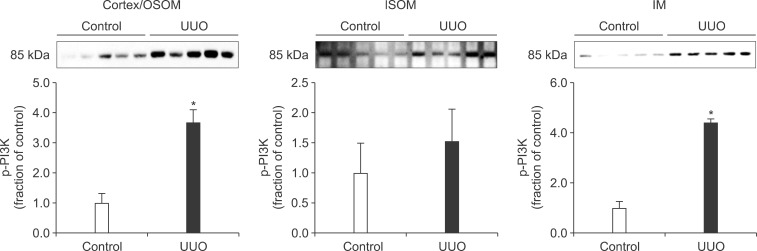
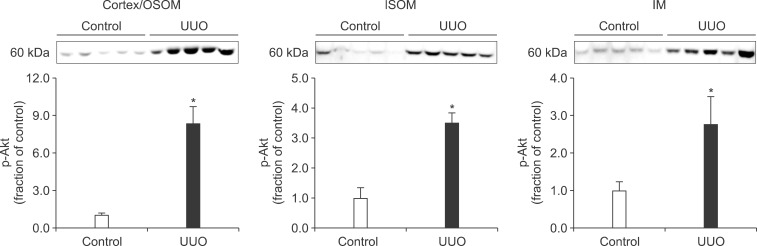
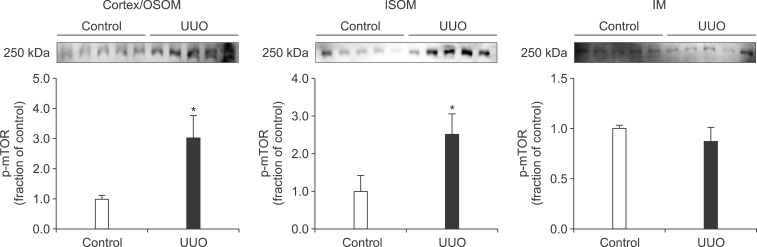
In the ureteral obstructed kidney, the expression of p-PI3K was significantly increased in the cortex/OSOM and IM, whereas it was not significantly changed in the ISOM (Fig. 1). The expression of p-Akt was also increased in the cortex/OSOM, ISOM, and IM (Fig. 2). The expression of p-mTOR was increased in the cortex/OSOM and ISOM (Fig. 3).
3. Expression of TGF-β1, Bax, and Bcl-2
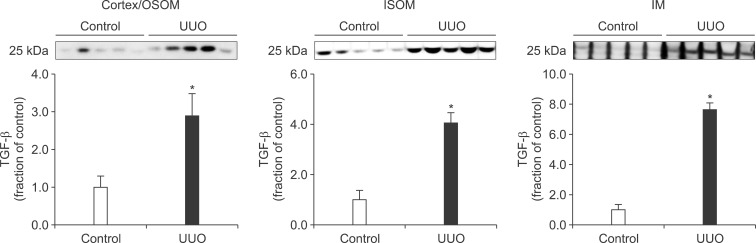
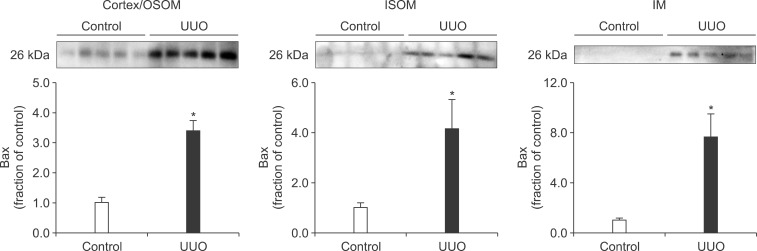
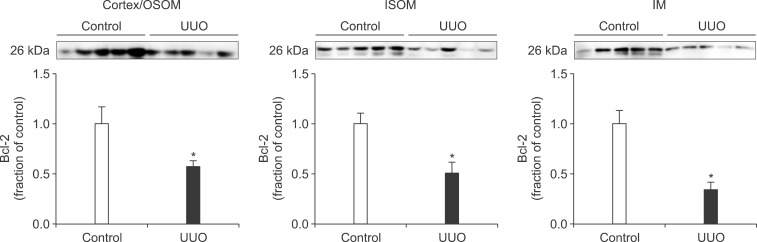
Following the UUO, the protein expression of TGF-β1 was increased in the cortex/OSOM, ISOM, and IM of the obstructed kidney (Fig. 4). The expression of Bax was also increased in the obstructed kidney compared with the control (Fig. 5). The protein expression of anti-apoptotic factor Bcl-2 was decreased in the obstructed kidney (Fig. 6).
DISCUSSION
Interstitial fibrosis and tubular atrophy are common consequences of chronic ureteral obstruction. TGF-β1 is considered to be a major stimulating factor that regulates extracellular matrix synthesis. It has been suggested that TGF-β1 is increased at the transcriptional level in chronic ureteral obstruction and that it may play a role in initiating fibrogenesis in obstructive nephropathy.12 In addition, it has been established that cellular apoptosis is associated with the upregulation of Bax and down-regulation of Bcl-2. In the present study, the protein expression of TGF-β1 was increased in the obstructed kidney following UUO. The expression of apoptotic factor Bax was increased in the obstructed kidney but that of anti-apoptotic factor Bcl-2 was decreased. These changes may contribute to the kidney injury in UUO. However, diverse mechanisms are involved in the regulation of TGF-β1, Bax, and Bcl-2, and the upstream mechanisms remain elusive.
mTOR is a serine/threonine kinase that is activated through the phosphorylation of PI3K and Akt.13,14 Recently, the mTOR signaling pathway has emerged as an important modulator of several forms of renal disease. The activation of mTOR in the kidney promotes the upregulation of pro-inflammatory and pro-fibrotic factors, which leads to tubulointerstitial fibrosis and atrophy.5-7 The expression of mTOR was increased in the kidney of DOCA-salt hypertensive rats, which was related to the upregulation of TGF-β1, pro-inflammatory proteins, and pro-apoptotic proteins.11 In addition, treatment with mTOR inhibitor ameliorates kidney injury in experimental diabetic nephropathy and a rat model of reduced renal mass through inhibition of TGF-β1 and proinflammatory cytokines.8,9,15,16 In the present study, the expression of p-PI3K, p-Akt, and p-mTOR was significantly increased in the ureteral obstructed kidney. These changes may contribute to the interstitial fibrosis and tubular atrophy in chronic ureteral obstruction.
In conclusion, the phosphorylation of PI3K/Akt/mTOR was increased in the obstructed kidney of rats with UUO, which may be related to the pathogenesis of tubulointerstitial fibrosis and apoptosis.
 XML Download
XML Download