

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Glaucoma is one of the leading causes of irreversible blindness.1 Glaucoma is an optic neuropathy characterized by retinal ganglion cell (RGC) death, axon loss, and an excavated appearance to the optic nerve (ON) head.2 In glaucoma, elevated intraocular pressure (IOP) and other factors, such as oxidative stress and interruption of axoplasmic transport, have been implicated in RGC death.3-8
Although elevated IOP is the most significant risk factor for glaucomatous ON damage and RGC loss, several studies have shown a relation between oxidative stress and RGC loss in glaucoma.8,9-11 In addition, there is much evidence of a dysregulated blood supply to the ON head and abnormal retinal blood flow in glaucoma patients and in a chronic ocular hypertensive animal model.12-16
Histopathologically, the loss of RGCs in glaucoma is accompanied by morphologic and functional changes in Müller cells the main type of glial cell in the retina.17 Müller cells are activated in the retina under stressful conditions, such as elevated IOP or ischemia.17-19 A key feature of activation of Müller cells is the upregulation of the intermediate filament glial fibrillary acidic protein (GFAP).20-23
Under normal conditions, Müller cells are responsible for the protection of RGCs by releasing neurotrophic factors and the secretion of glutathione, which has an antioxidant effect.24-26 However, activated Müller cells negatively affect RGC survival in ischemic retina.26-28
Melatonin was recently found to be an antioxidant and a free radical scavenger.29,30 It is a highly effective direct scavenger of reactive radicals and their intermediates, such as hydrogen peroxide, nitric oxide, and peroxynitrite. Because of its lipophilic characteristic, which allows it to cross the blood-brain barrier, melatonin has been proposed to endogenously protect against oxidative damage to the brain.31,32
In this study, we investigated the expression of hypoxia inducible factor 1α (HIF-1α) and GFAP in ischemic mouse retina. In addition, to determine the neuroprotective effect of melatonin, RCG counting by retrograde FluoroGold labeling was performed in ischemic retina treated with either vehicle or melatonin.
MATERIALS AND METHODS
1. Animals
All experiments were conducted in accordance with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research. The protocol was approved by the Institutional Animal Care and Use Committee of Chonnam National University Hospital. C57BL/6 mice (3 months of age, 20-25 g in weight) were housed in individual cages under controlled lighting conditions (12 hours light/12 hours dark) and given tap water and food ad libitum.
2. Transient retinal ischemia
For transient retinal ischemia, C57BL/6 mice were anesthetized with a mixture of tiletamine/zolazepam (4 mg/kg; Zoletil 50, Virbac, France) and xylazine hydrochloride (1 mg/kg, Rompun; Bayer Korea, Seoul, Korea) by intramuscular injection. A cannula was inserted into the anterior chamber, which was connected by flexible tubing to the saline reservoir. IOP was elevated above systolic blood pressure (100-120 mmHg) for 60 minutes by raising the reservoir. Retinal ischemia was confirmed by the whitening of the anterior segment of the eye, blanching of the iris vessels, and loss of retinal red reflex. Previous studies have shown that neuronal degeneration occurs mainly in the thinner retinal layer, especially in the ganglion cell layer, after 60 minutes of transient retinal ischemia.18,33
3. Melatonin treatment
Two groups of mice were studied following unilateral transient retinal ischemia: one group was treated with vehicle (saline, n=10) and the other group was treated with melatonin (40 mg/kg, n=10) 1 hour before ischemia, at the time of ischemia, and 1 hour after ischemia. The dose and delivery route of melatonin were based on previous reports for melatonin treatment.34
4. Tissue preparations
The retinas were dissected from the choroid and fixed in 4% paraformaldehyde in phosphate buffered saline (pH 7.4) for 2 hours at 4℃ for retinal flat mounting. For Western blot analyses, whole retinas were immediately used or frozen in liquid nitrogen and stored at -70℃ until used.
5. Western blot analysis
Retinal tissues were homogenized in a glass-polytetrafluoroethylene Potter homogenizer in lysis buffer (PRO-PREP™; iNtRoN Biotechnology, Kyungki-Do, Korea). Each sample (10 µg) was separated in a 10% polyacrylamide mini-gel at 150 V for 1 hour. The transferred membranes were incubated for 1 hour at room temperature in TBS-T solution [10 mM Tris-HCl (pH 7.6), 150 mM NaCl, and 0.1% Tween-20] containing 5% non-fat dry milk. Blocking membranes were incubated overnight at 4℃ with mouse monoclonal anti-HIF-1α (1:2000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), mouse monoclonal anti-GFAP antibody (1:3000; Cell Signaling Technology), or mouse polyclonal anti-actin antibody (1:4000; Santa Cruz Biotechnology) in TBS-T solution containing 5% non-fat dry milk. After being washed three times with TBS-T, the membranes were incubated for 1 hour at room temperature with peroxidase-conjugated goat anti-mouse IgG (1:2000; Santa Cruz Biotechnology) in TBS-T containing 5% non-fat dry milk. Blots were developed with enhanced chemiluminescence and quantified by using an LAS-3000 image-analyzer (Fujifilm, Tokyo, Japan).
6. Retrograde labeling and counting of RGCs
FluoroGold (1 µL/injection of 4%; Fluorochrome, Inc., Denver, CO, USA) diluted in saline was microinjected bilaterally into the superior colliculi of anesthetized mice in a stereotactic apparatus 1 week before enucleation. To evaluate the loss of RGCs, each retinal quadrant was divided into three zones: center, middle, and peripheral retina (one-sixth, three-sixths, and five-sixths of the retinal radius). The numbers of RGCs were counted in 32 distinct areas of 0.48 mm2 (two areas at the center and three areas at the middle and periphery per retinal quadrant) by two investigators in a masked fashion, and the scores were averaged. The images were analyzed by confocal microscopy with a 20× objective with a laser scanning microscope (LSM 510; Carl Zeiss, Jena, Germany).
RESULTS
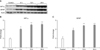
1. Expression of HIF-1α and GFAP on ischemic mouse retina according to time interval
Western blot showed increased expression of HIF-1α and GFAP in ischemic mouse retina within 24 hours after ischemia-reperfusion. Expression of HIF-1α and GFAP peaked 24 hours after ischemia-reperfusion (p<0.05; Fig. 1).
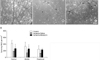
3. Effect of melatonin on RGC loss
Compared with the control retina, acute IOP elevation induced RGC loss by approximately 45% in the central, 58% in the middle, and 48% in the peripheral areas of vehicle-treated ischemic retina 2 weeks after ischemia-reperfusion (Fig. 4, A, B, and D; Table 1). In contrast, melatonin treatment increased RGC survival in ischemic mouse retina, especially in the central and middle areas compared with vehicle-treated ischemic retina (p<0.05; Fig. 4, C and D; Table 1).
DISCUSSION
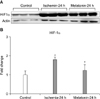
This study showed that melatonin treatment inhibited the increased expression of HIF-1α and GFAP and prevented RGC death in ischemic mouse retina. When adequate oxygen is supplied to the cell, HIF-1α is usually degraded in the cytoplasm. However, under hypoxic conditions, stabilized HIF-1α forms heterodimer with HIF-1β in the nucleus.35,36 This heterodimer transcribes several genes associated with cellular adaptation to hypoxia, such as erythropoietin, vascular endothelial growth factor, and nitric oxide.37
Therefore, modulation of HIF-1α is tightly regulated by the level of tissue hypoxia. However, excessive expression of HIF-1α can lead to neuronal cell death.38,39
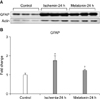
Our result showed that melatonin inhibits increased expression of HIF-1α in ischemic retina compared with ischemic retina treated with vehicle. We suggested that the possible mechanism of this inhibition might be mediated by the antioxidant effect of melatonin and that blockage of subsequent hypoxic responses via inhibition of HIF-1α expression results in increased RGC survival in ischemic retina. In addition, we investigated the effect of melatonin on GFAP expression in ischemic retina. Melatonin effectively reduced GFAP expression and increased RGC survival in ischemic retina. Müller cells, the principal glial cells of the retina, support neurons with blood-derived nutrients, by removing metabolic waste, and by maintaining the homeostasis of the retinal extracellular milieu (ions, water, neurotransmitter molecules, and pH).40 Müller cells become activated or reactive in response to pathological alteration of the retina, including ischemia-reperfusion or elevated IOP. Furthermore, activated Müller cells may participate in damaging neurons41,42 and are characterized by upregulation of the intermediate filament protein GFAP.43
However, the exact mechanism responsible for activation of Müller cells is still unknown. Activated Müller cells increase the expression of the inducible form of nitric oxide synthase and decrease glutamate uptake in ischemic retina.44,45 Tezel and Wax46 reported that increased production of tumor necrosis factor-α and nitric oxide by retinal glial cells exposed to elevated hydrostatic pressure and simulated ischemia induced apoptosis in cocultured RGCs.
Melatonin is an indoleamine produced in the pineal gland and the retina. Melatonin is known to be involved in diverse functions in the human body, such as temperature regulation, blood pressure regulation, and sleep-cycle control.47 In the retina, melatonin receptors are clustered on the photoreceptors, inner segments, and ganglion cells.46 Melatonin is known to inhibit the nitridergic pathway, which has a protective effect on the photoreceptor's outer membranes and reverses the effect of ocular hypertension on retinal function. In addition, in an experimental animal model, the concentration of melatonin in the retina of glaucomatous rats with high IOP was significantly reduced.48 Melatonin has diverse direct and indirect antioxidant effects and also acts as a free radical scavenger. Several reports have indicated the potent antioxidant and neuroprotective effects of melatonin. Treatment with melatonin after neonatal hypoxia-ischemia was shown to reduce neuronal cell death and white-matter demyelination in rats.49 In addition, injection of melatonin after induction of hypoxic-ischemic injury reduced oxidative stress and inflammatory cell recruitment in the cerebral cortex of rats.50 Moreover, melatonin counteracted ischemia-induced apoptosis in cultured human retinal pigment cells.51 The antioxidant functions of melatonin include direct free radical scavenging, stimulation of antioxidative enzymes, increased efficiency of mitochondrial oxidative phosphorylation, reduction in electron leakage, and augmentation of the efficiency of other antioxidants.52-56 Therefore, as a potent antioxidant, melatonin can induce functional changes in Müller cells during ischemia-reperfusion injury. We believe that the decreased activation of Müller cells resulting from melatonin treatment can reduce RGC death during ischemia-reperfusion injury. Thus, the use of melatonin may be an effective therapeutic strategy for preventing glaucomatous RGC death. In combination with the effects of traditional anti-glaucoma medication, the antioxidative effect of melatonin may benefit patients with glaucoma by protecting against RGC death resulting from ischemic injury. In conclusion, our findings provide evidence that the functional state of retinal glial cells may be important in determining the ultimate fate of RGCs and that melatonin has a neuroprotective effect on RGC in ischemic retina, and this effect is mainly mediated by a reduction in the stabilization of HIF-1α and activation of Müller cells. The specific molecular events that occur in Müller cells after melatonin treatment need to be determined.

 XML Download
XML Download