

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Sphingosine-1-phosphate (S1P) is derived from sphingosine, the backbone of most sphingolipids, and is now emerging as a vital lipid mediator.1 S1P is one of a new class of second messengers involved in cellular proliferation, differentiation, and apoptosis and implicated in diverse physiological functions, including immune modulation, vascular and nervous system development, regulation of smooth muscle, and auditory and vestibular function.1-4
Intermediates in the biosynthesis and catabolism of sphingolipids, ceramide, sphingosine, and S1P have recently been implicated in the intracellular signaling important for neuronal survival, differentiation, development, and death. Because increased levels of ceramide were observed during differentiation of neuroblastoma Neuro2a cells and SH-SY5Y cells, it has been suggested that ceramide plays a role in neuronal differentiation.2,5,6 Also, S1P was recently shown to be a key regulator of proliferation and differentiation by up-regulating sphingosine kinase expression in retina photoreceptors.7
Neuroblastomas derived from immature sympathetic ganglionic cells are arrested at various stages of differentiation.8 Differentiation studies have revealed that neuroblastoma cell lines can be induced to differentiate in the presence of various agents and growth factors. At first, human SH-SY5Y neuroblastoma cells (a subclone of the SKN-SH cell line) were reported to differentiate morphologically and biochemically in response to bioactive phorbolesters.9
Despite many studies concerning the physiologic actions of S1P, little is known about its capacity to differentiate neurons. This study intended to explore the differentiating action of S1P by using neuroblastoma cell lines. In this study, S1P induced neuronal differentiation in neuroblasoma cells.
MATERIALS AND METHODS
1. Cell culture and treatment
SH-SY5Y human neuroblastoma cells (ATCC number: CRL-2266™) capable of differentiation to neuronal cells under specific conditions were cultured in DMEM (Hyclone, Logan, UT, USA) containing 10% fetal bovine serum (Hyclone) and 1% penicillin-streptomycin (Hyclone). The cells were grown to approximately 80% confluence in a 37℃ humidified incubator in an environment of 5% CO2 and 95% air and were then harvested in trypsin containing EDTA (Hyclone). Sphingosine-1-phosphate (Sigma Chemical Co., St. Louis, MO, USA) was initially dissolved and diluted to a final stock concentration of 10 mM in water before use. The SH-SY5Y neuroblastoma cells were differentiated for 3 or 4 days in the presence of 1 or 10 uM S1P.
2. Analysis of neurite outgrowth
The cells were grown under conditions with S1P for 4 days. The morphology of the SH-SY5Y cells was investigated with a NiKon phase-contrast inverted microscope equipped with a Nikon Coolpix 4500 high-resolution camera. Changes in neurite length were observed over 4 days, and the medium was replaced every 2 days. A total of 200 to 300 cells were microscopically evaluated and scored for neurite formation by use of the Image J program if they had a neurite that was longer than one cell diameter or had a growth cone. All experiments were repeated at least three times with similar results.
3. Reverse transcription-polymerase chain reaction (RT-PCR)
Cells were cultured in 6-well plates as described previously. Total RNA was extracted from the cultured cells by using the Tri Reagent (Molecular Research Center Inc., Cincinnati, OH, USA) isolation reagent. cDNA was synthesized by reverse transcription with M-MLV reverse transcriptase (Gibco BRL, Grand Island, NY, USA) and 1 mmol total RNA. The cDNA was amplified by 25 to 35 cycles of PCR (Takara Bio Inc., Shiga, Japan) with Ex-Taq polymerase (Takara Bio Inc.). The primers (Bioneer Co., Daejeon, Korea) used were chosen and checked for specificity by using a basic BLAST search. The RT-PCR products were separated electrophoretically on 2% agarose gels. We used the following primer pairs (gene, forward, reverse, and cDNA product length): 5'-AAA TCG CAT CCA GAT TTT C-3', 5'-CAC TGC CTC CTA GCT TGT C-3', 316 bp; neurofilament-H (NFH), 5'-CCG ACG ACA CCA AGC TCA-3', 5'-GGA ATG AAA CAG GGC GTT-3', 336 bp; microtubule-associated protein-2 (MAP-2), 5'-TGC CAT CTT GGT GCC GA-3', 5'-CTT GAC ATT ACC ACC TCC AGG T-3', 460 bp; synaptophysin (SYP), 5'-CTT CCT GCA GAA CAA GTA CC-3', 5'-CTT AAA CAC GAA CCA CAG GT-3', 295 bp; neurogenic differentiation 1 (NeuroD1), 5'-CCG ACA GAG CCC AGA TGT AGT TCT T-3', 5'-GCC CCA GGG TTA TGA GAC TAT CAC T-3', 278 bp; synaptotagamin (SYT), 5'-CGC AAA CTG GGC AAA CGC TA-3', 5'-GCA ACC CTC GTG GGC CTC-3', 421 bp; GAPDH, 5'-CAT GAC CAC AGT CCA TGC CAT CAC T-3', 5'-TGA GGT CCA CCA CCC TGT TGC TGT A-3', and 461 bp.
4. Immunostaining
Immunochemical determination of neuronal differentiated SH-SY5Y cells was performed as follows. The cells were grown on poly-D-lysine-coated aclar plastic coverslips for 4 days in the presence of 10 mM S1P, fixed for 15 min with 4% paraformaldehyde (Sigma Chemical Co.), permeabilized for 20 min with 0.3% Triton X-100 (Sigma Chemical Co.) including 10% normal goat serum (Vector Laboratories Inc., Burlingame, CA, USA) in phosphate-buffered saline, and incubated with antibodies specific for each antigen marker. The cultures incubated with primary antibodies were followed by biotinylated secondary antibodies and avidin-biotin complex (Vector Laboratories Inc.). The cell type-specific markers used were growth associated protein-43 (GAP43), NFH, and SYP. The antibodies were purchased from Chemicon (Chemicon, Temecula, CA, USA).
RESULTS
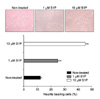
1. Effects of S1P on neurite outgrowth in neuroblastoma cells
After incubation with 1 uM or 10 uM S1P, the neuroblastoma cells revealed neurite outgrowth. The number of neurite-bearing cells increased from 14% to 37% and 28% by 1 uM S1P after 72 hours and 96 hours, respectively. The number of neurite-bearing cells increased from 14% to 18% and 46% by 10 uM S1P after 72 hours and 96 hours, respectively (Fig. 1).
2. Effects of S1P on the immunoreactivity for neuron-specific markers in neuroblastoma cells
After incubation with S1P, the neuroblasoma cells revealed immunoreactivity for neural-specific markers. The neuronal cells were positive for neuronal markers such as GAP43, NFH, and SYP (Fig. 2).
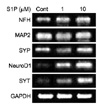
3. Effects of S1P on the expression of neuronal gene markers
Reverse transcription-polymerase chain reaction was used to examine the expression of mRNA related to neuronal differentiation. The expression of NFH, Ngn1, MAP2, SYP, NeuroD1, and SYT mRNA, which is specific for neurons, was increased by treatment with S1P (Fig. 3).
DISCUSSION
Neuronal differentiation and cellular homeostasis are fundamental events in the development of the nervous system as well as in the regeneration of damaged nervous tissue. Differentiation is regulated by a complex mechanism, in which the result of several interactions between the cell and the extracellular medium determines the fate of the cellular function.10
S1P is known to induce differentiations in various cell types. It stimulates differentiation of adipose tissue-derived mesenchymal stem cells towards smooth muscle cells. It increases the expression of smooth muscle cell-specific proteins such as alpha-smooth muscle actin (alpha SMA) and transgelin.11 Sphingosine kinase, which promotes the formation of S1P, enhances the expression of myogenic differentiation markers in myoblasts.12 Recently, Miranda et al proposed S1P as a key regulator in the development of photoreceptors. It increases the proliferation of photoreceptor progenitors and increases the formation of apical processes and enhances the expression of opsin and peripherin in photoreceptors.13
Despite their tumoral origin, neuroblastoma cell lines can be induced to differentiate in vitro by several agents.14 In this work, we used the SH-SY5Y neuroblastoma cell line, which showed homogeneous populations of differentiated cells due to STP or retinoic acid.15 Retinoic acids are natural and synthetic derivatives of vitamin A that, together with their nuclear control genetic programs, are essential for embryonic development, organ homeostasis, cell growth, differentiation, and apoptosis.16
We observed that treatment with S1P for 4 days induced neuronal morphological changes in neuroblastoma cells, which were similar to those described by other authors.15,16 To confirm that the morphological changes observed actually resulted from the neuronal differentiation, the expression of several neuronal markers was explored. We observed that the typical morphological feature of differentiation was accompanied by biochemical changes. In the present study, S1P induced immunoreactivities for neuronal markers and increased the expression of neuronal gene markers in neuroblastoma cells.
Intermediates in the biosynthesis and catabolism of sphingolipids, ceramide, sphingosine, and S1P have recently been implicated in intracellular signaling important for neuronal survival, differentiation, development, and death. S1P is emerging as a new class of second messenger involved in cellular proliferation, differentiation, and apoptosis.2 The above experimental results imply that S1P could be used to differentiate neuronal cells and may be a good candidate for the treatment of neurodegenerative diseases.

 XML Download
XML Download