

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The nuclear envelope is a physical barrier which regulates the traffic between nucleoplasm and cytoplasm. It is a phospholipid bilayer membrane which consists of two layers; inner and outer membrane [12]. Inner and outer membranes are separated by the perinuclear space. The cytoplasm is connected to the nucleoplasm via nuclear pores. Although small size of molecules (less than 30 kDa) freely move through the nuclear pore, bigger molecules need the help of special carrier proteins. In the nuclear pore, the nuclear pore complex (NPC) limits the transportation of macromolecules including protein or RNAs.
Recently, new roles of the nuclear pore in gene expression, chromatin organization and DNA repair have been reported in addition to its roles as a physical barrier. Moreover, its abnormal changes like mutation, copy number alteration and mislocalization of molecules, have been associated with various pathologic conditions such as cancers, genetic disorders, and neurodegeneration [23456]. So, investigating its mechanism and disease-associated changes should be helpful for developing novel therapeutic strategies. These abnormal changes could be potential drug targets. Moreover, the necessity to deliver therapeutic DNA or proteins into the nucleus has arisen to treat diseases such as cancer and genetic diseases. Recent progresses in the research of the molecular mechanism for the nuclear transport via NPC, factors affecting the nuclear transport and the application for therapeutics will be summarized in this review.
Nuclear Transport Cycle
Transportation of macromolecules including protein or RNAs between nucleoplasm and cytoplasm occurs through NPC in the nuclear envelope. NPC is highly selective and bidirectional transporter for various cargo molecules. There are four important factors for the nuclear transport: (1) nucleoporins (NUPs) that are constituent proteins of NPCs, (2) RanGTPase that allows for active transport and directionality, (3) karyopherins (importin/exportin/transportin) that can recognize cargo molecules, (4) nuclear localization signals (NLSs) or nuclear export signals (NESs) in cargo molecules. NLSs or NESs are recognized by karyopherins. After briefly reviewing the overall process of nuclear transport cycle, four factors will be discussed.
Import cycle
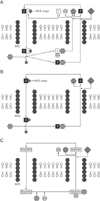
First step of the nuclear transport cycle is the formation of importin-cargo complex. Importins can bind cargo molecules after recognition of their NLSs. Depending on types of NLSs, different importins are involved. For example, classical NLSs can be recognized by importin-α in the cytoplasm where RanGTP is very rare. Then, the N-terminus (importin-β binding domain) of importin-α binds to importin-β (Fig. 1A) [27]. However, importin-β can directly recognize proline-tyrosine (PY) NLS without importin-α, and this pathway is fast and efficient (Fig. 1B) [789101112].
After the formation of importin(s)-cargo complex, importin-β is specifically recruited to NPC in the nuclear pore and then the complex can pass through the nuclear pore. In the nucleoplasm, binding of RanGTP to importin-cargo complex facilitates dissociation of cargo, and then RanGTP-bound importin is exported to the cytoplasm. Recycling of importin-α is mediated by a nuclear export receptor, cellular apoptosis susceptibility protein (CAS), but CAS is not necessary to export of importin-β (Fig. 1).
Export cycle
Cargos containing NES are recognized by exportins. The NES-bound exportin forms a complex with RanGTP. RanGTP is 100-fold more abundant in the nucleoplasm than in the cytoplasm. Binding of RanGTP to exportins induces the high affinity of exportin for its cargo molecule [213]. The exportin-cargo-RanGTP complex specifically binds the docking site of NPCs and then passes through nuclear pore. RanGAP in the cytoplasm hydrolyzes RanGTP in the complex, which leads to the dissociation of the complex (Fig. 1) [21314]. RanGDP is reimported into the nucleoplasm with help of nuclear transport factor 2 [1315].
Factors Affecting Nuclear Transport
Nucleoporins
NPC is a complex basket-like structure with huge molecular mass of 120 MDa in humans. It made up of NUPs with 100–150 nm in diameter and 50–70 nm in thickness according to the species [161718]. Each NPC has a central hole (~30 nm in diameter and ~50 nm in long) that connects between the nucleoplasm and the cytoplasm [17]. Main structures of NPC include the inner pore ring, the nuclear and cytoplasmic rings, the nuclear basket, and the cytoplasmic basket. NPC is composed of multiple copies of 30–50 different NUPs and 500–1,000 NUPs are integrated into the NPC structure. They are functionally conserved throughout eukaryotic cells [2171920].
NUPs serve as an architectural scaffold and a permeability regulator. Some NUPs have intrinsically disordered domains rich in repeating amino acid sequences such as FXFG (Phe-X-Phe-Gly), Phe-Gly (FG), or GLFG (Gly-Leu-Phe-Gly), which act as docking sites for karyopherin-βs (importin or exportin) [161819202122232425262728293031]. These repeating sequences line the central hole and regulate the passage of cargo molecules. NUP62 complex which consists of NUP62, NUP58, and NUP54 has been shown to include FG-repeats [32].
Ran
RanGTP gives the nuclear transport directionality. The association of RanGTP with exportins moves the complex out of the nucleoplasm. However, its association with importins leads to the dissociation with cargo molecules. RanGTP is about 100-fold more abundant in the nucleoplasm than in the cytoplasm, which is possible due to the fact that Ran's guanine-exchange factor RCC1 is located in the nucleoplasm but Ran GTPase activating protein, Ran GAP in the cytoplasm [213].
Karyopherin: importin and exportin
More than 20 karyoperins in human have been reported. Among them, 11 karyopherins (importin-β, importin-β2, importin-4, importin-5, importin-7, importin-8, importin-9, importin-11, transportin-SR, importin-13, exportin-4) are involved in import or bidirectional transport of cargo molecules. Eight exportins (exportin-1–exportin-7, exportin-t) are involved in the export of cargo molecules. Each karyopherin has its own specific cargo molecules. For example, CAS (exportin-2) contributes to the export of importin-α and exportin-t to the export of t-RNA [14].
Importin-β have high molecular flexibility because it can have structural changes using their tandem Huntingtin, elongation factor 3 (EF3), protein phosphatase 2A (PP2A), and the yeast kinase TOR1 (HEAT) repeats which can be thought as a helical spring [17333435]. A single HEAT motif consists of a pair of α-helices and both helices are amphiphilic. Because of an unusual hydrophobic core that supports intramolecular helix-helix interactions, the molecular structure of importin-β is highly flexible [153637]. CAS has similar structure to importin-β and it is based on 19 HEAT repeats [1537].
Importin-α has been shown to contain a tandem series of Armadillo (ARM) repeats that are composed of an array of Trp, Asn, and acidic residues on the inner surface [2383940]. Although ARM motif has three helical structures instead of two in HEAT motif, the superhelical 3D structure of ARM repeats is similar with that of HEAT repeats.
Karyopherins can also export nucleotide motif including tRNA, miRNA, rRNA, viral RNA, and uridine-rich small nuclear RNAs with or without ribonucleoprotein as an adaptor [71441424344]. However, structure of mRNAs is highly diverse unlike other RNAs, so that they can be transported by processing and assembly into mRNs [14].
NLS and NES
NLS
The best well characterized NLS is classical NLSs for the nuclear protein import. Classical NLSs contain monopartite or bipartite signals that can be recognized by importin-α. A first characterized classical monopartite NLS is simian virus 40 (SV40) large T antigen (PKKKRKV) (Fig. 2). Bipartite NLSs have two clusters of basic amino acids sequence. The prototypical bipartite NLS is nucleoplasmin found in Xenopus laevis (KRPAATKKAGQAKKKK) (Fig. 2) [431344546474849]. The classical NLS is thought as the typical NLS because it was the first NLS to be defined.
In contrast to classical NLSs, PY-NLS has diverse sequence and large structure, nonetheless, importin-βs can recognize them by multiple attractions of weak affinities between NLS and importin [72125]. PY-NLS sequences are composed of a loose N-terminal hydrophobic motifs and a C-terminal RX2-5PY motif [735]. hnRNP A1, Hrp1 are the representative PY-NLSs (Fig. 2) [3550].
Karyopherin121 (Kap121) is one of the most essential for nuclear transport in Saccharomyces cerevisiae and it can mediate transportation for diverse cargos [5152]. Recent studies demonstrated that the small lysine-rich NLSs (consensus sequences: K-V/I-X-K-X1-2-K/H/R) interact with Kap121 [5152].
Transportin3 (Trn3) binds the cargo containing RS (Arg-Ser) repeats domain especially phosphorylated RS repeats. In proteomic analysis, about 32% of Trn3 cargos have RS repeats [5153]. Other cargos of Trn3 contain RE (Arg-Glu) or RD (Arg-Asp) motifs, which may imitate phosphoRS motifs [51]. According to identification of new cargos for importins, additional consensus sequences for new classification will be required.
There are many other NLSs recognized by importin-α. Representative sequences from Borna Disease Virus p10 protein and S. cerevisiae phospholipid scramblase 1 contain hydrophobic residues (Fig. 2) [1754]. Importin-β also binds various other NLSs included in CREB, ribosomal proteins, the human immunodeficiency virus Rev and Tat, the human T-cell leukemia virus type 1 protein Rex, PTHrP, cyclin B1, Smad3, SREBP-2, and TRF (Fig. 2) [3789101112133536375055565758].
NES
The consensus sequence for NESs is Φ1-X(2-3)-Φ2-X(2-3)-Φ3-X-Φ4 motif (Φ: represents hydrophobic residues L, I, F, M, or V and X: any amino acid). Different exportins have their specific cargo molecules. For example, CAS (exportin-2) transports importin-α. CRM1 (exportin-1) is a ubiquitous nuclear export receptor containing hydrophobic residues. Binding site of CRM1 consists of five pockets [113161823242730]. CRM1 can recognize relatively diverse molecules. Overexpression of CRM1 was noted in many types of cancer [21920].
Structure of NLS-bound complex
Several factors such as NLSs, NESs, and 3D structures need to be considered to improve the efficiency of nuclear transport. NLSs are well-studied part in the nuclear transport, and commercial NLS peptides including HIV-Tat, penetratin, and (Arg)9, are available for the gene delivery. However, as the importance of conformation is being emphasized, many researches have focused on the 3D structures.
NLSs can be recognized as linear and/or conformational signals by importins. The 3D structure of NLSs would be changed in NLSs-bound complex, which causes changes of binding affinity with importin. Many groups has made many efforts to bridge the gap between 3D structure and binding capacity of complex (Fig. 3) [41424344]. Karyopherins have different binding sites according to their subtypes and binding residues of certain karyopherin can be different according to different kinds of cargo. One study demonstrated that the C-terminal and N-terminal structures of NLS significantly affect the efficiency of the nuclear transport as well as their binding affinity to importin [5960]. Kim et al. [61], compared with structurally modified SV40 NLS peptides. They made modified peptides from SV40 NLS by addition of cysteine, deletion of cysteine, homodimerization, or circularization. These modifications showed different transfection efficiency according to structures even though the NLS has same sequences.
Furthermore, although many researchers have tried different amount of NLS peptides which were fused with cargos and different locations for the nuclear delivery, the effects of them still require further study [596061]. Overall, alteration of 3D structure can change the binding affinity to importin and the conformation is a more important factor to be considered when we use NLS peptides for the nuclear delivery.
Clinical Significance of Nuclear Transport
In multiple cancers, changes in the expression of karyopherins including importin-α2, CAS, importin-β1, exportin-1, have been reported [24662]. Overexpression of exportin-1 has been associated with various cancers including acute and chronic leukemia, multiple myeloma, ovarian cancer, pancreatic cancer, gastric cancer, sarcoma, melanoma, glioma, and cervical cancer [62]. A small molecule inhibitor for exportin-1 showed some beneficial effects in the clinical trial [62]. In cancer cells, exportin-1 played critical roles in exporting various tumor suppressor proteins including retinoblastoma protein, adenomatous polyposis coli, p53, p21, p27, FOXO, IκB, topoisomerase II, and PAR-4 [26].
Furthermore, abnormal localization of proteins due to abnormalities in the nuclear transport, can cause various disorders that involve protein aggregation, biosynthesis, or cell metabolism [4663646566676869]. For example, sex-determining region Y (SRY) is transported by a specific importin to the nucleus to activate testis-related genes. There are two NLS motifs in the DNA-binding domain of SRY that are called high-mobility group box. Mutations of NLS of SRY have been observed to be associated with Swyer syndrome. Swyer syndrome is a sex reversal disease of male caused by XY gonadal dysgenesis [464548]. A similar alteration was reported in NLS of trichorhinophalangeal syndrome type 1 gene in patients with trichorhinophalangeal syndrome type I [669]. Additionally, alterations in several NLSs of disease-related proteins have been identified; short stature homeobox in patients with Léri-Weill dyschondrosteosis, aristaless related homeobox in patients with X-linked lissencephaly, forkhead box P2 in patients with speech-language disorder [6656667].
Therapeutic Applications of NLS
The necessity to deliver therapeutic DNAs or proteins into the nucleus has been increased to correct genetic or nuclear abnormalities. Although its efficiency is high, viral transduction showed several critical problems such as immunogenic response and safety risk. Non-viral methods are safe but their delivery efficiency are low [70].
To increase the efficiency of the nuclear delivery, NLSs have been used. Instead of long sequences of NLSs, researchers inserted short NLSs like SV40, (Arg)x or HIV-tat. Some studies developed a plasmid DNA containing the NLS of SV40, and thereby the transfection efficiency of the target DNA was increased [6870]. The insertion of NLS showed great results for the delivery of CRISPR/CAS system. For example, Ramakrishna et al. [71], developed (Arg)9-mediated RNA-guided endonucleases (RGEN) delivery process [71727374]. Furthermore, TAT-based gene delivery systems was also developed to enhance the efficiency of nuclear import [75]. Simple insertion of NLSs in the target molecules cannot be the solution for delivery to the nucleus, because NLSs can be hindered in inserted status. So, 3D structure of drugs should be considered for the nuclear targeting.
Many researchers have tried to develop ideal drug delivery systems by using modification of NLSs [72737677]. Based on previous results, several bioinformatic tools, such as NucPred, NLS Mapper, NESbase and NLSdb, have been developed about subcellular localization of proteins [787980818283]. Using these tools, we can search the subcellular locations of targeting molecules. More importantly, users can predict subcellular location of their own drugs containing NLSs by using NucPred website (http://www.sbc.su.se/~maccallr/nucpred/) [78]. Fig. 2 shows the predictive subcellular location of popular NLS sequences based on NucPred.
Based on its clinical significance, further studies about roles and molecular mechanisms about the nuclear transport need to be done in the future study. How to improve the efficiency of NLSs for the nuclear delivery has been discussed. To enhance the delivery efficiency by using NLSs, several factors such as its types and 3D structures should be considered. If we can predict nuclear transport efficiency based on 3D structures of cargos-karyopherin complex, it might be very helpful to develop treatment methods.


 XML Download
XML Download