

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Transient global cerebral ischemia (tGCI), which is caused by lack of blood supply to the brain, results in irreversible neuronal death in venerable regions of the brain, such as the hippocampus and cerebral cortex [12]. The hippocampus is extremely sensitive to cerebral ischemic injury [3]. In particular, pyramidal neurons in the cornu ammonis 1 (CA1) are killed from several days after tGCI [45].
Medicinal plants, which have been used to heal various ailments in humans, have received considerable interest in research fields for treatments of various diseases including neurological disorders, because many medicinal plants have a variety of biological properties [678]. The genus Populus (poplar) belonging to the Salicaceae family includes various species distributes in template zone and subtropical areas, and several species of genus Populus have been used in traditional medicine [910]. For example, Populus nigra and Populus davidiana possess pharmacological actions including antioxidant, anti-inflammatory and hepatoprotective properties [1112]. Furthermore, Populus balsamifera has pharmacological potential for treatment of obesity and diabetes [13]. However, among Populus species, few studies regarding scientific validation of pharmacological actions of P. tomentiglandulosa (Korea poplar), which is a species of the genus Populus, have been reported. Especially, to the best of our knowledge, protective effects of P. tomentiglandulosa against cerebral ischemic insults have never been investigated. Therefore, in the present study, we examined neuroprotective effects of P. tomentiglandulosa extract in the hippocampal CA1 using gerbils, which are a good model of tGCI [1415].
Materials and Methods
Preparation of Populus tomentiglandulosa extract
P. tomentiglandulosa was collected in September 2014 by Professor Jong Dai Kim in Kangwon province (South Korea). For the preparation of ethanol P. tomentiglandulosa extract (PTE), P. tomentiglandulosa was washed with distilled water, air-dried at 60℃ and ground into fine powder by a grinder (IKA M20, IKA, Staufen, Germany). The P. tomentiglandulosa powder was refluxed with 10 vol (v/w) of 70% ethanol at 70℃ for 24 hours, and the extraction procedure was repeated three times. The extract was filtered via Whatman No. 1 filter paper (Whatman Ltd., Maidstone, Kent, UK), concentrated with a vacuum evaporator, and it was completely dried with a freeze-drier. The extraction yield was 14.5%.
Pre-treatment with PTE
We used male Mongolian gerbils (6 months of age and 70–80 g of body weight). Experimental protocol was approved (approval No. KW-160802-1) by the Institutional Animal Care and Use Committee at Kangwon National University. Animals were divided into 6 groups (n=7 in each group): (1) vehicle-sham group, which was treated with vehicle (sterile normal saline; 0.9% w/v NaCl) and given sham operation, (2) vehicle-tGCI group, which was treated with vehicle and given tGCI, (3) and (4) PTE-sham group, which was treated with 100 and 200 mg/kg PTE, respectively, and given sham operation, and (5) and (6) PTE-tGCI group, which was treated with 100 and 200 mg/kg PTE, respectively, and given tGCI. PTE was dissolved in saline, and PTE or saline was orally administered once a day for 7 days before ischemic surgery. We adapted the oral administration of PTE for 1 week, because extracts from plants have been taken orally in traditional medicine and no data regarding the absorption and metabolism of PTE haven reported.
Induction of tGCI
In brief, according to our published method [4], the gerbils were anesthetized with a mixture of 2.5% isoflurane in 33% oxygen and 67% nitrous oxide, and both common carotid arteries were occluded for 5 minutes. Restoration of blood flow (reperfusion) was directly observed under an ophthalmoscope. Body (rectal) temperature was maintained under normothermic (37±0.5℃) condition before, during and after tGCI. Sham-operated animals were subjected to the same surgical procedures except that the common carotid arteries were not occluded.
Tissue processing for histology
As we described previously [15], briefly, the gerbils were anesthetized with sodium pentobarbital (30 mg/kg, intraperitoneally) and perfused transcardially with 4% paraformaldehyde 5 days after tGCI. Their brains were removed and postfxed in the same fxative for 6 hours, cryoprotected by infiltration with 30% sucrose, and the frozen bran tissues were serially sectioned into 30-µm coronal sections in a cryostat (Leica, Wetzler, Germany).
Cresyl violet staining
To examine distribution pattern of cells in the ischemic hippocampus, the hippocampal sections were stained with cresyl violet (CV) according to our protocol [15]. Briefly, cresyl violet acetate (Sigma-Aldrich, St. Louis, MO, USA) was dissolved at 1.0% (w/v) in distilled water, and glacial acetic acid was added to this solution. The sections were stained, subsequently dehydrated by immersing in serial ethanol baths and mounted with Canada balsam (Tokyo, Japan).
Immunohistochemistry
In order to examine neuronal damage and change in astrocytes and microglia, immunohistochemistry was carried out according to our published method [16]. Briefly, we used mouse anti-neuronal nuclear antigen (NeuN, 1:1,000, Chemicon, Temecula, CA, USA) for neurons, mouse anti-glial fibrillary acidic protein (GFAP, 1:800, Chemicon) for astrocytes, and rabbit anti-ionized calcium-binding adapter molecule 1 (Iba-1, 1:800, Chemicon) for microglia as primary antibodies. The sections were incubated with each primary antibody, exposed to biotinylated goat anti-mouse or rabbit IgG and streptavidin peroxidase complex (Vector Laboratories, Burlingame, CA, USA), and visualized with 3,3′-diaminobenzidine solution.
Data analysis
Data analysis were done as previously described [16]. In brief, for quantitative analysis of NeuN-immunoreactive (+) neurons, we selected 10 section with 150-µm interval in each animal, and captured digital images of the CA1 with an AxioM1 light microscope (Carl Zeiss, Jena, Germany) equipped with a digital camera (Axiocam, Carl Zeiss) connected to a PC monitor. NeuN+ neurons were counted in a 250×250 µm square applied at the center of the CA1 using an image analyzing system (software: Optimas 6.5, CyberMetrics, Scottsdale, AZ, USA). Cell counts were obtained by averaging total cell numbers.
For quantitative analyses of GFAP+ and Iba-1+ structures, their images were calibrated into an array of 512×512 pixels corresponding to a tissue area of 140×140 µm (40× primary magnification). Each immunoreactivity was measured by 0–255 gray scale system, and a ratio of relative optical density (ROD) for GFAP+ and Iba-1+ structures was calibrated as % using Adobe Photoshop version 8.0 and then analyzed using NIH Image 1.59 software. A ratio of the ROD was calibrated as %, with the vehicle-sham group designated as 100%.
Statistical analysis
The data shown here represent the means±SEM. Differences of the means among the groups were statistically analyzed by one-way analysis of variance (ANOVA) with a post hoc Bonferroni's multiple comparison test in order to elucidate ischemia-related differences among experimental groups. Statistical significance was considered at P<0.05.
Results
Neuroprotective effects
CV-positive (CV+) cells
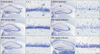
In the vehicle-sham group, cells were well stained with CV in all layers of the hippocampus (Fig. 1A, B). In the 100 mg/kg and 200 mg/kg PTE-sham groups, the distribution pattern of CV+ cells was similar to that in the vehicle-sham group (Fig. 1E, F, I, J).
In the vehicle-tGCI group, pyramidal cells in the stratum pyramidale were not stained with CV only in the CA1 5 days after tGCI (Fig. 1C, D). In the 100 mg/kg PTE-tGCI group, the CA1 pyramidal cells were not stained with CV 5 days after tGCI, which means that CA1 pyramidal cells were not protected from tGCI (Fig. 1G, H). On the other hand, in the 200 mg/kg PTE-tGCI group, the CA1 pyramidal neurons were well stained with CV 5 days after tGCI, which means that CA1 pyramidal cells were well protected from tGCI (Fig. 1K, L).
NeuN+ cells
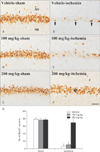
The CA1 pyramidal cells showed strong NeuN immunoreactivity in all sham operated groups (Fig. 2A–C), and no significant difference in the number of NeuN+ cells between those groups was observed (Fig. 2G).
In the vehicle-tGCI group, a few NeuN+ cells (7±2.3 neurons) were found in the CA1 stratum pyramidale at 5 days post-tGCI (Fig. 2D, G). In the 100 mg/kg PTE-tGCI group, the distribution and number of NeuN+ cells (11±3.5 neurons) were not significantly different from those in the vehicle-tGCI group (Fig. 2E, G). However, in the 200 mg/kg PTE-tGCI group, many NeuN+ cells (70±2.7 neurons) were detected in the CA1 stratum pyramidale, and the mean number of NeuN+ cells was similar to that in the vehicle-sham group (Fig. 2F, G).
Effects on glial activation
GFAP+ astrocytes
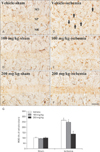
In the vehicle-sham group, normal GFAP+ astrocytes were found in the strata oriens and radiatum of the CA1, and they showed small cell bodies and thread-like processes (Fig. 3A). In the 100 and 200 mg/kg PTE-sham groups, the morphology, distribution and ROD of GFAP+ astrocytes were similar to those in the vehicle-sham group (Fig. 3B, C, G).
In the vehicle-tGCI group, the cytoplasm of GFAP+ astrocytes was hypertrophied with thickened processes, and the ROD of GFAP+ structures was significantly increased 5 days after tGCI compared with that in the vehicle-sham group (about 264% of the vehicle-sham group) (Fig. 3D, F). In the 100 mg/kg PTE-tGCI group, the morphology and ROD of GFAP+ astrocytes were not significantly different from those in the vehicle-tGCI group (Fig. 3E, G). Whereas, in the 200 mg/kg PTE-tGCI group, GFAP+ astrocytes were significantly less hypertrophied (Fig. 3F), and their ROD was significantly deceased compared with those in the vehicle-tGCI group (about 122% of the vehicle-sham group) (Fig. 3G).
Iba-1+ microglia
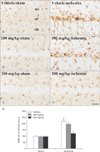
In the vehicle-sham group, Iba-1+ microglia showed small cytoplasm and long processes, and they were distributed throughout the CA1 (Fig. 4A). In the 100 and 200 mg/kg PTE-sham groups, the morphology and distribution of Iba-1+ microglia and their ROD were not changed compared with those in the vehicle-sham group (Fig. 4B, C, G).
In the vehicle-tGCI group, Iba-1+ microglia showed large cytoplasm and thick processes, and many of them were aggregated in the stratum pyramidale (Fig. 4D). In addition, in this group, the ROD of Iba-1+ microglia was significantly increased compared with that in the vehicle-sham group (about 220% of the vehicle-sham group) (Fig. 4G). In the 100 mg/kg PTE-tGCI group, the morphology, distribution and ROD of Iba-1+ microglia were not significantly different from the vehicle-tGCI group (Fig. 4E, G). However, in the 200 mg/kg PTE-tGCI group, their morphology and distribution were not significantly changed from those in the vehicle-sham group (Fig. 4F), and their ROD was significantly low compared with that in the vehicle-tGCI group (about 122% of the vehicle-sham group) (Fig. 4G).
Discussion
Until now, numerous researchers have tried to find medicinal plants which have pharmacological potentials for therapeutic applications in cerebral ischemia, and many studies in vivo and in vitro have reported that extracts from various medicinal plants and their components show therapeutic effects against cerebral ischemia [81718]. However, therapeutic effects of PTE against tGCI has not yet been investigated. Therefore, in the present study, we examined neuroprotective effects of PTE in the gerbil hippocampal CA1 subjected to 5 minutes of tGCI and found that many of pyramidal neurons survived in the ischemic CA1 of gerbils pre-treated with 200 mg/kg PTE when we examined the CA1 pyramidal cells using CV staining and NeuN immunohistochemistry, which are universally used staining techniques for examining neuronal death in the brain [119]. This finding is the first in proving neuroprotective effects of the P. tomentiglandulosa against tGCI.
It has been well demonstrated that cerebral ischemia causes reactive gliosis of astrocytes and microglia and that reactive astrocytes and microglia are involved in cerebral ischemia-induced neuronal death through production and release of neurotoxic molecules and pro-inflammatory cytokines [2021]. Therefore, it has been thought that inhibiting reactive astrocytes and microglia is considered to protect neurons from cerebral ischemic insults [2223]. In this study, we examined reactive gliosis of astrocytes and microglia in the ischemic CA1 after pretreatment with PTE via GFAP and Iba-1 immunohistochemistry, respectively. Immunohistochemical results showed that tGCI-induced reactive gliosis of astrocytes and microglia was significantly diminished by pretreatment with 200 mg/kg of PTE. Although there were no studies regarding effects of Populus species on the reactive gliosis induced by cerebral ischemic insults, it was reported that extracts of some species such as P. davidiana and Populus maximowiczii strongly inhibited production of nitric oxide, which is considered as a pro-inflammatory mediator, in lipopolysaccharides (LPS)-stimulated macrophages [24]. In addition, a recent study demonstrated that salicortin isolated from Populus euramericana extract significantly suppressed expressions of pro-inflammatory cytokines, such as tumor necrosis factor-α, interleukin (IL)-1β and IL-6 in macrophages stimulated by LPS [25]. Therefore, it is likely that the inhibition of tGCI-induced reactive gliosis in the ischemic CA1 by PTE pretreatment might be closely associated with neuroprotection of PTE against tGCI.
In brief, the results of this study obviously show that pretreatment with PTE can protect CA1 pyramidal neurons from ischemic injury following tGCI and that its neuroprotective effect is closely related to attenuation of reactive gliosis. Therefore, our recent study suggests that P. tomentiglandulosa can be a promising candidate for prevention of cerebral ischemia-induced neuronal damage/death, although further researches are positively necessary to explain neuroprotective mechanisms of P. tomentiglandulosa against tGCI.
 XML Download
XML Download