

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Allergic diseases of the respiratory system such as asthma and rhinitis are a significant health concern in developing countries due to increased environmental pollution [1]. The rapid growth in the prevalence, severity, and complexity of allergic respiratory diseases in modern population imposes a significant burden on quality of life and health-care costs [2]. The close association between mast cell and early-phase of hypersensitivity plays a vital role in allergic diseases [34]. The specific antigen such as FcεRI activates the mast cells that are present in all organs and tissues [56]. The IgE/FcεRI-mediated activation causes release of various chemical mediators in early-phase (such as histamine, β-hexosaminidase [β-HEX], proteoglycans, proteases, and tryptase) as well as in late-phase (proinflammatory cytokines such as tumor necrosis factor α [TNF-α], interleukin [IL]-1β, IL-4, IL-5, IL-6, IL-13, transforming growth factor-β, and vascular endothelial growth factor). These mediators play an important role in induction and maintenance of immune-inflammatory reactions [7].
An array of studies over past decade confirmed compound 48/80 (C48/80) as a potent and well-established agent for mast cell degranulation and to evaluate the mechanism of new therapeutic moieties against allergic reactions [89]. C48/80 can rapidly release inflammatory substances including histamine from rat peritoneal mast cells (RPMCs) [10]. This allergic inflammatory influx results in various pathophysiologic events including vasodilation, increase vascular permeability, tissue edema, bronchial and intestinal smooth muscle contraction, and elevated mucus production [11]. These conditions contributes to various disease state including allergic hyper-responsiveness, migraine [12], pain [1314], interstitial cystitis [15], irritable bowel syndrome [16], and endometriosis [17].
Over past decade, disodium cromoglycate (DSCG) has been used extensively as a “mast cell stabilizer” and serves as a treatment option for management of various allergic diseases including asthma, allergic rhinitis, allergic conjunctivitis, and mastocytosis [1819]. However, use of DSCG for the mast cell stabilization is limited because of its nephrotoxicity [1920]. Therefore, search for novel agents derived from plant material for mast cell stabilization potential is ongoing. Researchers reported therapeutic potential of many isolated moieties from herbal extracts such as curcumin [9], caffeic acid [21], resveratrol [22], and epigallocatechin gallate [23] against mast cell degranulation. Furthermore, polyphenols isolated from apple extract was reported for the inhibitory potential against mast cell degranulation via inhibition of binding between FcεRI and IgE [242526].
Type-A proanthocyanidins polyphenols isolated from cinnamon (Cinnamomum zeylanicum Blume, Family: Lauraceae) bark (TAPP-CZ) shown to have an array of therapeutic applications for the treatment of immune inflammation, arthritis, asthma, and viral infection [2728293031]. Recently, the anti-allergic potential of TAPP-CZ against ovalbumin-induced experimental allergic rhinitis has been reported in mice after its intranasal administration [32]. However, the molecular mechanism(s) of action of TAPP-CZ for the management of allergic responses remains unresolved. Hence, the present study aimed at the mast cell stabilization as a possible mechanism for the anti-allergic action of TAPP-CZ against the mast cell degranulation induced by C48/80 on isolated RPMCs.
Materials and Methods
Animals
Sprague-Dawley rats (180–200 g) were purchased from National Toxicology Centre, Pune and kept in quarantine for 1 week. Experimental protocol (SIOP/IAEC/2011/22) was approved by the Institutional Animal Ethics Committee of Sinhgad Institute of Pharmacy, Pune, India. The rats were maitained in the animal house of the institute under standard laboratory conditions (temperature of 24±1℃, relative humidity of 45%–55% and 12:12 hour light/dark cycle) throught the study. Animals had free access to standard chow pelleted food (Pranav Agro-industries Ltd., Sangli, India) and water ad libitum. Animals were brought to the testing laboratory 1 hour for adaptation purpose before the experiments timings (between 10:00 AM to 5:00 PM).
Chemicals
C48/80, MTT, and Percoll were purchased from Sigma Chemical Co. (St. Louis, MO, USA). DSCG was purchased from Ahlcon Parenterals (India) Ltd. (New Delhi, India). All other reagents were purchased from SD Fine Chemicals, Mumbai, India. Enzyme-linked immunosorbent assay (ELISA) kits for histamine (Cat No. BA D-0024-10499 from Labor Diagnostika Nord GmbH & Co. KG, Nordhorn, Germany), IL-4 (Cat No. ELR-IL4-001C from RayBiotech Inc., Norcross, GA, USA), and β-HEX (Cat No. E90637Ra from USCN Life Science Inc., GmbH, Germany) were purchased from respective suppliers.
Test compound
The test compound, TAPP-CZ, prepared as per reported procedure [3233], was provided by Indus Biotech Private Limited (Pune, India). TAPP-CZ was characterized by high-performance liquid chromatography fingerprinting (Jasco, Tokyo, Japan) and found to match with characteristics of TAPP-CZ from cinnamon bark as reported earlier [3435]. The TAPP-CZ contains pentameric type-A procyanidins flavonoid (75.9% purity) as a marker compound with dimer (1.7%), trimer (11.1%), and tetramers (2.266%). The test compound complies with quality requirements related to the absence of microbial content and heavy metals.
Preparation of RPMCs
The RPMCs were isolated as per previously reported procedure [36]. In brief, rats were anesthetized with ether, injected with 10 ml of calcium-free hydroxyethyl piperazineethanesulfonic acid (HEPES)-Tyrode buffer into the peritoneal cavity, and the abdomen gently massaged for approximately 90 seconds. The peritoneal cavity was opened and the fluid aspirated using a Pasteur pipette. RPMCs were purified using a Percoll density gradient as per reported procedure [37]. RPMC preparation was approximately 95% pure as assessed by toluidine blue staining [38]. Purified RPMCs (1×106 cells/ml) were resuspended in HEPES-Tyrode buffer.
RPMCs viability assay
The viability of RPMCs was tested by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay as per reported method [39]. Briefly, RPMCs (2×105 cells/well) were pretreated with various concentrations of either vehicle (distilled water, 10 µg/ml), DSCG (10 and 100 µg/ml), or TAPP-CZ (1, 3, 10, and 30 µg/ml) at 37℃ for 3 hours. After addition of MTT (5 mg/ml saline), RPMCs was incubated at 37℃ for 4 hours. After washing out the supernatant, the insoluble formazan the crystallized MTT was dissolved in dimethyl sulfoxide and the absorbance was measured at 570 nm with a spectrophotometer (UV-visible spectrophotometer, V-630, Jasco). The density of formazan formed by vehicle RPMCs was taken as 100% of viability.
Mast cell stabilization activity
Cell viability was checked by a trypan blue dye (0.4%) exclusion test [38]. RPMCs (2×105 cells/well) were pretreated with various concentrations of either DSCG (10 and 100 µg/ml) or TAPP-CZ (1, 3, 10, and 30 µg/ml) or vehicle (10 µg/ml) at 37℃ for 3 hours and was incubated with C48/80 (1 µg/ml) at 37℃ for 10 minutes. The mast cell numbers (total and degranulated) were counted under high power microscope (Motic digital microscope, Motic Asia-Pacific, Hong Kong) for a set of 100 cells per treatment group. The % intact cells were calculated by the formula: Percentage of intact mast cells=(Total number of mast cells−Total number of degranulated cells)/(Total number of mast cells)×100.
Histamine release assay
The RPMCs suspensions (2×105 cells/ml) were pretreated with various concentrations of either DSCG (10 and 100 µg/ml) or TAPP-CZ (1, 3, 10, and 30 µg/ml) or vehicle (10 µg/ml) at 37℃ for 3 hours and incubated with C48/80 (0.25 µg/ml) for 15 minutes. The cells were separated from the released histamine by centrifugation (at 400 ×g for 5 minutes at 4℃). Residual histamine in the cells was released by disrupting the cells with perchloric acid and centrifugation at 400 ×g for 5 minutes at 4℃. The histamine content was measured by using histamine ELISA assay kit on a microplate reader at 450 nm.
β-HEX release assay
The β-HEX assay was performed as described previously with some modifications [40]. The RPMCs suspensions (2×105 cells/ml) were pretreated with various concentrations of either DSCG (10 and 100 µg/ml) or TAPP-CZ (1, 3, 10, and 30 µg/ml) or vehicle (10 µg/ml) at 37℃ for 3 hours and incubated with C48/80 (0.25 µg/ml) for 15 minutes. The 100 µl of supernatant was used for the β-HEX enzyme ELISA. Briefly, 100 µl of supernatant was added to β-HEX substrate buffer (100 mM sodium citrate, 1 mM 4-nitrophenyl N-acetyl-β-D-galactosaminide, pH 4.5) and incubated for 1 hour at 37℃. The reaction was terminated using stop solution, and absorbance was measured using a microplate reader at 450 nm.
IL-4 release assay
The RPMCs suspensions (2×105 cells/ml) were pretreated with various concentrations of either DSCG (10 and 100 µg/ml) or TAPP-CZ (1, 3, 10, and 30 µg/ml) or vehicle (10 µg/ml) at 37℃ for 3 hours and incubated with C48/80 (0.25 µg/ml) for 15 minutes. The 100 µl of supernatant was used for IL-4 sandwich ELISA as per manufacturer's instructions (RayBiotech, Inc.) using a microplate reader at 450 nm.
Statistical analysis
The data was presented as a mean±standard error of mean (SEM) and analyzed by one-way analysis of variance (ANOVA) using GraphPad Prism 6.0 software (GraphPad, San Diego, CA, USA). Dunnett's test was applied for post hoc analysis. The P-values less than 0.05 (two-sided) were considered significant.
Results
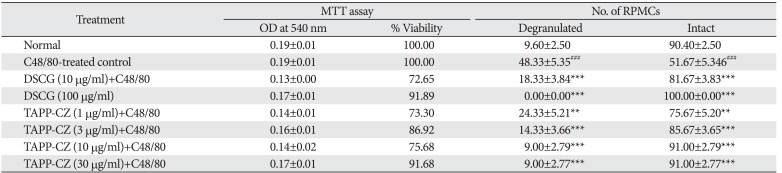
Effect of TAPP-CZ on RPMCs viability assay
The cell viability remains 100% in TAPP-CZ (3 and 30 µg/ml) and DSCG (100 µg/ml) treated RPMCs during MTT assay. The % viability of the RPMCs after pretreatment with TAPP-CZ (1 and 10 µg/ml) as well as DSCG (10 µg/ml) was found to be around 80% (Table 1).
Effect of TAPP-CZ on mast cell stabilization activity
The C48/80 treated control RPMCs showed significant (P<0.001) increase in the number of degranulated cell and decrease in intact cells as compared to normal RPMCs. The DSCG pretreated (10 and 100 µg/ml) and TAPP-CZ pretreated (3, 10, and 30 µg/ml) RPMCs showed significant (P<0.001) reduction in the number of degranulated cells as compared with C48/80-treated control RPMCs (Table 1).
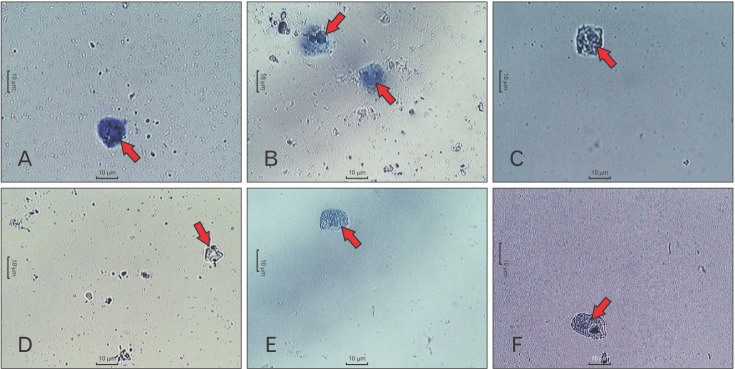
Effect of C48/80 on mast cell morphology
Normal RPMCs were oval and contained many fine granules surrounding a prominent nucleus (Fig. 1A). After stimulation with C48/80, RPMCs were degranulated (characterized by cell swelling) and showed the presence of extruded granules near the cell surface (Fig. 1B). When RPMCs were pretreated with DSCG (100 µg/ml) and TAPP-CZ (3, 10, and 30 µg/ml), they showed inhibition of C48/80-induced degranulation (Fig. 1C–F, respectively).
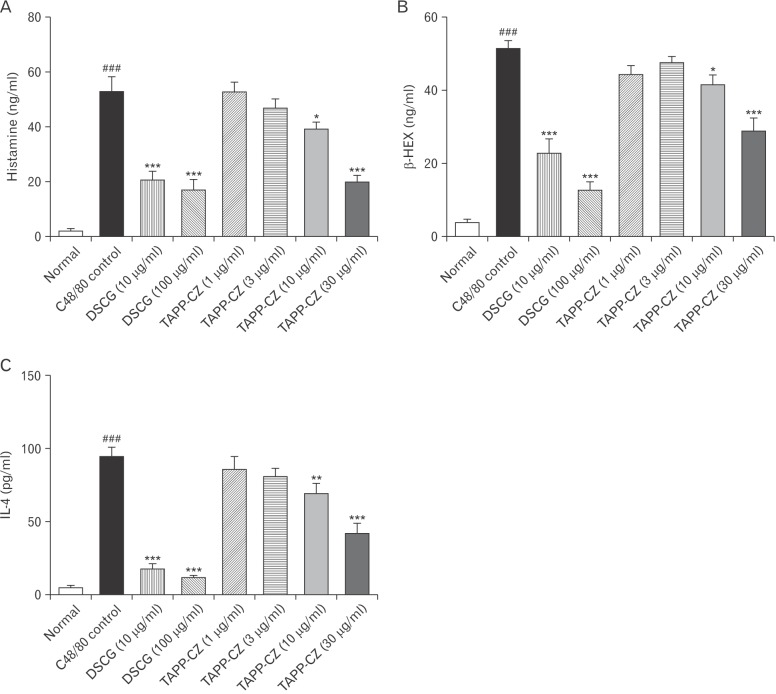
Effect of TAPP-CZ on C48/80-induced histamine release in RPMCs
A significant increase (P<0.001) in the level of histamine in C48/80-treated control RPMCs were found as compared to normal RPMCs. Pretreatment with TAPP-CZ (10 and 30 µg/ml) showed the significant attenuation (P<0.05 and P<0.001, respectively) of elevated histamine levels as compared to C48/80-treated control RPMCs. Pretreatment with DSCG (10 and 100 µg/ml) showed significant decrease (P<0.001) in histamine the levels as compared to C48/80-treated control RPMCs (Fig. 2A). The half maximal inhibitory concentration (IC50) of TAPP-CZ for histamine release from RPMCs was found to be 1.48 µg/ml.
Effect of TAPP-CZ on C48/80-induced β-HEX release in RPMCs
The β-HEX levels of C48/80-treated control RPMCs were significantly (P<0.001) more than normal RPMCs. TAPP-CZ (10 and 30 µg/ml) treated RPMCs showed significant and dose dependant attenuation (P<0.05 and P<0.001) in β-HEX levels as compared to C48/80-treated control RPMCs. DSCG (10 and 100 µg/ml)-treated RPMCs also showed the significant (P<0.001) decrease in the elevated levels of β-HEX levels as compared to C48/80-treated control RPMCs (Fig. 2B).
Effect of TAPP-CZ on C48/80-induced IL-4 release in RPMCs
A significant increase (P<0.001) in the level of IL-4 was found in C48/80-treated control RPMCs as compared to normal RPMCs. The significant and dose-dependent attenuation (P<0.01 and P<0.001) of IL-4 level was found in TAPP-CZ (10 and 30 µg/ml)-treated RPMCs as compared to C48/80-treated control RPMCs. The elevated level of IL-4 was significantly (P<0.001) inhibited by the pretreatment of DSCG (10 and 100 µg/ml) when compared with control RPMCs (Fig. 2C). The IC50 of TAPP-CZ for IL-4 release from RPMCs was found to be 24.44 µg/ml.
Discussion
Mast cell plays a significant role in acute and chronic allergic inflammation after FcεRI stimulation by releasing various proinflammatory mediators such as histamine, cytokines, chemokines, and leukotrienes [4142]. Allergy is known as type I hypersensitive or early-phase of reaction to various allergens occurred in response to activation of mast cells and basophils via interaction with IgE, resulting in immune-inflammatory responses [114143]. The mast cell contains high-affinity receptor, FcεRI, on its surface with an ability of antigen-specific IgE binding [4144]. C48/80 is the condensed product of N-methoxy phenylamine with formaldehyde, which is the most potent mast cell degranulator amongst the various secretagogues [45]. C48/80 has an ability to induce membrane perturbation that increases the permeability of the lipid bilayer membrane [46]. Moreover, its interaction with mast cell caused the release of almost 90% of histamine as compared to natural process. Therefore, in the present investigation, we used C48/80 to evaluate the mechanisms of action of TAPP-CZ against RPMCs. Furthermore, pre-treatment of RPMCs with TAPP-CZ retains its normal morphology, showed significant inhibition in a number of degranulated cell and histamine, IL-4 as well as β-HEX release from RPMCs.
In allergic patients, allergen challenge causes an early-phase reaction within an hour which is followed by a late-phase reaction after 3 to 48 hours [47]. A compelling body of evidence supports that the early-phase reaction which is an acute inflammatory process is characterized by the release of histamine and other pharmacological mediators from mast cells [4849]. Whereas, the release of pro-inflammatory cytokines such as TNF-α and IL's in various tissues including lung, nose, skin, and retina is a hallmark for the late phase reaction which is the clinical manifestation of chronic allergic diseases [5051]. Mast cells are involved in both early and late phase reaction in IgE-mediated allergic reactions.
The mast cell serves as a storehouse for histamine which is synthesis from its precursor L-histidine via activation of enzyme histidine decarboxylase. The release of histamine from activated mast cell results in proinflammatory response as well as modulation of the activity of a variety of cells including T cells, monocytes, macrophages, neutrophils and eosinophils [52535455]. These mediators are responsible for induction and maintenance of acute inflammation and early-phase of hypersensitivity [56]. Thus, mast cells played a central dogma role in early-type hypersensitivity and allergic diseases. In the present investigation, RPMCs stimulated with C48/80 resulted in mast cell degranulation thus elevates the levels of histamine whereas pretreatment with TAPP-CZ attenuated the elevated levels of histamine which might be due to inhibition of mast cells degranulation. Results of the present investigation are in line with the findings of the previous investigator where polyphenol treatment significantly inhibited histamine release from RPMCs [25].
The enzyme β-HEX is considered as the major mediator of the acute inflammation and early hypersensitivity responses along with histamine. It has been reported that interaction of β-HEX with cell-surface receptors resulted in the development of various symptoms such as acute rhinitis and bronchoconstriction [57]. Thus, controlling the release of histamine and β-HEX from mast cells is thought to be important targets for attenuation of various allergic conditions [4158]. Interestingly, in the present study, we found a similar result where incubation of C48/80 with RPMCs induced the release of β-HEX whereas pretreatment with TAPP-CZ showed inhibition of elevated β-HEX levels.
Recently, some researchers have reported that late-phase reaction is more accurate clinical features of disease as compared to early-phase response, as it involved pro-inflammatory mediators along with various types of cells [259]. During the late-phase reaction, the eosinophils are recruited after IgE-mediated stimulation of mast cell degranulation [60]. IL-4 is reported to act as an eosinophil chemoattractant that enables endothelial cells to produce eosinophil chemotactic factor and eotaxin [61]. It also plays an important role in conversion of naïve T cells to allergic Th2 cells. Thus, IL-4-dependent recruitment of eosinophil responsible for elevated inflammation during allergic diseases [62]. Clinically, it has been shown that patients with allergic rhinitis are associated with high serum IL-4 levels as compared to healthy controls [63]. These findings depicted the importance of inhibition of pro-inflammatory cytokines release from mast cell in amelioration of allergic inflammation. Our results of mast cell degranulation by C48/80 with elevated IL-4 levels are in agreement with the existing literature [6264]. However, pretreatment with TAPP-CZ inhibits mast cell degranulation and subsequently decreases the level of IL-4 to induce mast-cell-mediated allergic reactions.
Research revealed that allergic reactions could be significantly suppressed through attenuating the interaction between IgE and FcεRI [6566]. Numerous scientific studies also demonstrated that the compound with an ability to inhibit the binding of IgE antibody and FcεRI possesses strong anti-allergic action [8924]. The procyanidin polyphenol-enriched apple extract is reported to inhibit mast cell degranulation in-vitro via decreasing the intracellular cross-linking of IgE and FcεRI thus suggesting its antiallergenic potential [2467]. Furthermore, piceatannol, a stilbene-type polyphenol, was shown to inhibit mast cell degranulation and secretion of various mediators via decreasing IgE and FcεRI interaction [6869]. Another naturally occurring polyphenol compound, chlorogenic acid, was shown to inhibit mast cell-dependent anaphylactic reaction [70]. In the past, anti-allergic effects of TAPP-CZ against ovalbumin-induced allergic rhinitis in mice was shown to be mediated by serum IgE level inhibition [32]. Therefore, mast cell stabilization activity of TAPP-CZ can be attributed, at least partly, to the IgE inhibition.
In conclusion, the TAPP-CZ obtained from cinnamon bark demonstrated the beneficial effect against C48/80 induced of mast cell degranulation, probably due to inhibition of the cross-linking of IgE with FcεRI and reduces the levels of the allergic makers such as histamine, β-HEX, and IL-4.
 XML Download
XML Download