

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The suboccipital muscles are located in the deepest layer of the neck, comprising the four paired muscles of the rectus capitis posterior major (RCPma), the rectus capitis posterior minor (RCPmi), the obliquus capitis superior (OCS), and the obliquus capitis inferior (OCI) muscles. Unilateral contraction of these muscles results in head rotation, whereas bilateral contraction results in head extension [1]. An association between the suboccipital muscles and headache has recently been pointed out, and McPartland et al. [2] reported that atrophy of the suboccipital muscles following whiplash is involved in marked, chronic neck pain and reduced standing balance. Malformation, defects, and anomalies of the suboccipital muscles, including the presence of accessory muscles, might also affect head and neck pain [3456]. In Japan, Mori [7] have reported variations of the suboccipital muscles, but few studies have examined these muscle groups morphologically, and therefore many morphological aspects of these muscles remain unclear.
Some studies have shed light on the muscle fiber characteristics of skeletal muscles using cell biology techniques [8910]. Myosin heavy chain (MHC) is a contractile protein with the largest mass of all such proteins, and is known to directly reflect the characteristics of muscles [11]. Depending on their rate of contraction, muscle fibers are classified as fast-twitch, intermediate, and slow-twitch types [12]. Sartorius et al. [13] reported that skeletal muscles in myosin heavy chain-IId-null mice show increased levels of myosin heavy chain-IIa (MHC-IIa). Yanagisawa et al. [14] have also reported that the presence of MHC-IIa after weaning in microphthalmic (mi/mi) mice with no tooth eruption was attributable to compensation for a lack of proper masticatory function and sucking-like movements, as MHC-IIa is necessary for these movements. In other words, muscle fibers change according to muscle function and structure, and therefore an understanding of the composition of MHC is important when considering the function of the suboccipital muscles.
The present study investigated: (1) morphological variations in the suboccipital muscles, and (2) the muscle fiber characteristics of these muscle groups. On the basis of our findings, we also discussed the role of the suboccipital muscles.
Materials and Methods
This study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000) and was approved by the Human Research Ethics Committee at the dental school. We dissected 25 cadavers donated for the annual student dissection courses at Tokyo Dental College (14 males, 11 females; mean age 80 years at death; range, 53–97 years). The cause of death in each case had been ischemic heart failure. All cadavers had been donated to Tokyo Dental College for research and education on human anatomy, and their use for research did not require approval by the university ethics committee.
Morphological observation
Forty-four muscle specimens (22 cadavers) had been fixed by intravenous injection of non-neutralized 10% (v/v) formalin solution and preserved in 50% ethanol solution for more than 3 months. Morphological observation of the suboccipital muscles for variations in the muscle belly was performed. The RCPma, RCPmi, OCS, and OCI were dissected under a magnifying glass after resecting the trapezius, splenius capitis and semispinalis capitis muscles. Variations in the muscle bellies of the suboccipital muscles were classified into different types by macroscopic observation.
Immunohistochemistry
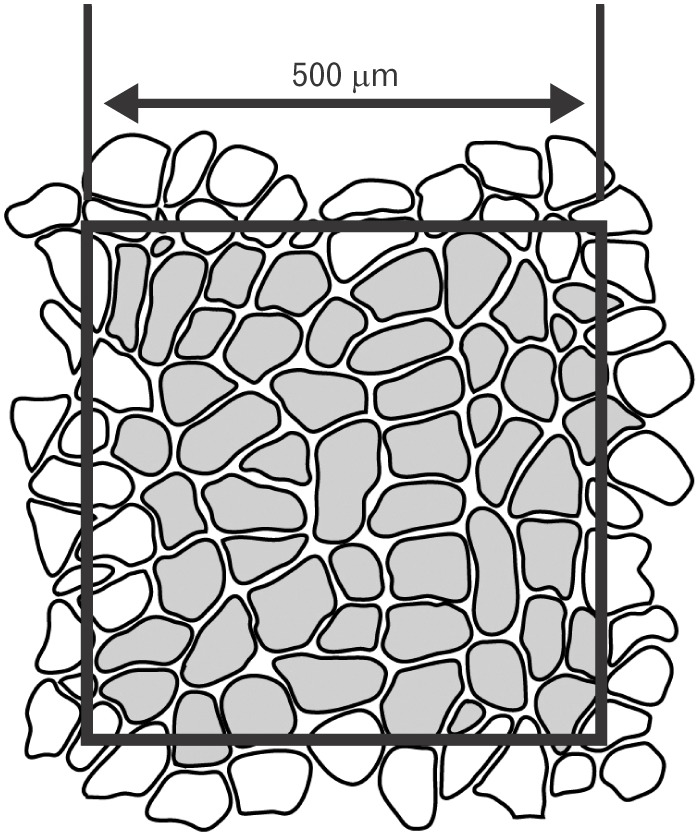
The remaining six muscle specimens (3 cadavers) were those of unfixed fresh cadavers, all of the individuals having died within the previous 3 days. These specimens were chosen as being representative of normal morphology because they showed no deformation or damage to the cervical spinal joints and no positional anomalies in the surrounding muscles beyond the area of observation. A square tissue slice measuring approximately 10 mm on all sides was harvested from the vicinity of the insertion of the suboccipital muscles and mounted on a piece of cork to ensure that the direction of the muscle fiber bundles was vertical. The tissue samples were then rapidly frozen in isopentane solution (−160℃) cooled with liquid nitrogen, and thereafter cryopreserved at −80℃ until use in experiments. Samples were prepared using a CM 3000 cryostat (Leica Microsystems, Wetzlar, Germany) to create 10-µm-thick serial sections on a plane perpendicular to the muscle fibers. The obtained serial sections were then subjected to immunostaining using mouse monoclonal anti-myosin heavy chain fast (MHCf) antibody (Abcam plc., Cambridge, UK) and mouse monoclonal anti-myosin heavy chain slow (MHCs) antibody (Sigma-Aldrich, St. Louis, MO, USA) as the primary antibody, and fluorescein isothiocyanate-donkey anti-mouse IgG antibody (Jackson Immuno Research, West Grove, PA, USA) as the secondary antibody. Slides were observed using an Axiophot 2 polarizing light microscope (Carl Zeiss, Jena, Germany) and the approximate center of the tissue sections was observed and photographed at ×100 magnification. Immunohistochemical examinations were performed using the method described by Doi et al. [9] for objective evaluation. Muscle fibers stained with anti-MHCf antibodies were designated as fast-twitch muscle fibers, those stained with anti-MHCs antibodies as slow-twitch muscle fibers, and those showing positive staining with both antibodies as intermediate fibers. Mean cross-sectional area (CSA) of muscle fibers per square 500 mm was determined using Image-Pro Plus software (Media Cybernetics, Rockville, MD, USA) (Fig. 1).
Results
Gross anatomical observations
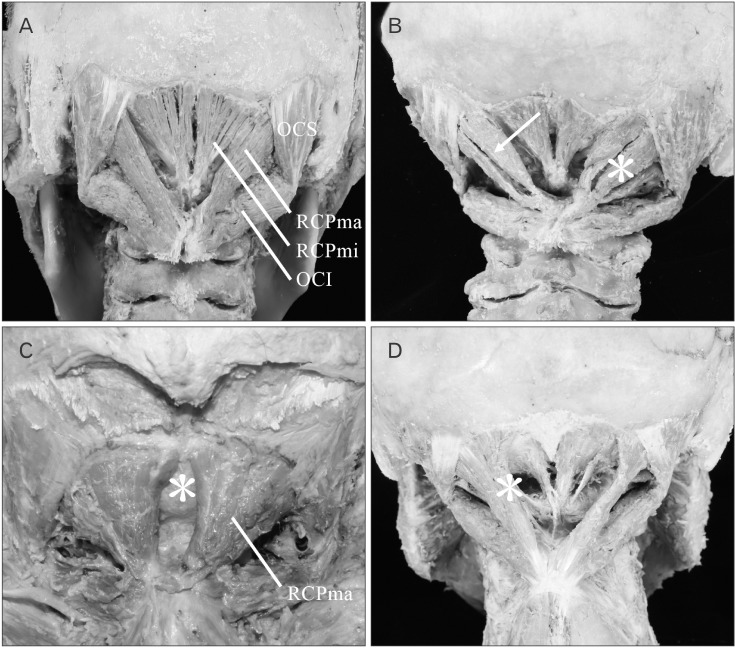
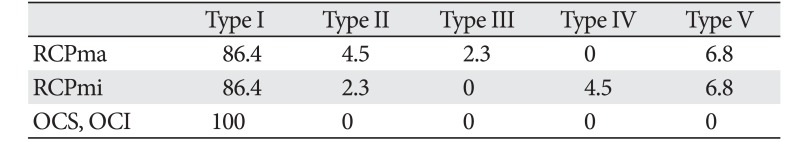
Macroscopic morphological observation revealed the following points of origin and insertion of each of the suboccipital muscles. The RCPma showed a tendinous origin from the spinous process of the second cervical vertebra and a tendinous insertion into the inferior nuchal line of the occipital bone. The RCPmi had a tendinous origin from the posterior tubercle of the first cervical vertebra and a tendinous insertion into the inferior nuchal line close to the occipital bone midline. The OCS displayed a tendinous origin from the transverse process of the first cervical vertebra and a tendinous insertion into the distal end of the inferior nuchal line of the occipital bone. The OCI had a tendinous origin from the spinous process of the second cervical vertebra and a tendinous insertion into the transverse process of the first cervical vertebra. As anomalous anatomy was evident for both the RCPmi and the RCPma, these muscles were classified into five types.
(1) Type I: The muscle is formed of muscle fiber bundles exhibiting a typical form of attachment and all muscle bellies form a single belly. Muscle fiber bundles have a tendinous origin and insertion (Fig. 2A) The RCPma and RCPmi were of this type in 38/44 (86.4%) and 38/44 (86.4%) of specimens, respectively (Table 1).
(2) Type II: All the muscle at the origin is consistent with typical muscle (Type I), but the insertion consists of a tendinous attachment to two adjacent locations, and two muscle bellies are present (Fig. 2B). A tendency for a wider area of attachment than the insertion of type I muscle is evident. The RCPma and RCPmi were of this type in 2/44 (4.5%) and 1/44 (2.3%) of specimens, respectively (Table 1).
(3) Type III: The origin is at the spinous process of the second cervical vertebra in the RCPma and at the posterior tubercle of the first cervical vertebra in the RCPmi. The insertion is at the inferior nuchal line of the occipital bone in both the RCPma and the RCPmi. While all muscle at the origin is consistent with typical muscle, the insertion consists of tendinous attachments to three adjacent locations, and three muscle bellies are present (Fig. 2B). A tendency for a wider area of attachment than the insertions of types I and II muscle is also evident. Type III muscle was seen in 1/44 (2.3%) of RCPma samples, but not in any of the RCPmi samples (Table 1).
(4) Type IV: The muscle belly is completely missing and the muscle is filled with fat tissue in its place (Fig. 2C). Two (4.5%) of the 44 RCPmi samples were classified as this type. Bilateral insertions of the RCPma in type IV are attached closer to the midline than in other samples, and the width of the muscle belly is greater. No defects of the muscle belly were seen in the RCPma (Table 1).
(5) Type V: Fusion of the muscle bellies is seen between the RCPma and RCPmi. The origin of the fused muscle bellies is at the posterior tubercle of the first cervical vertebra, matching the origin of the RCPmi. The insertion is at the inferior nuchal line of the occipital bone, consistent with the insertion of the RCPma (Fig. 2D). The RCPma and RCPmi were of this muscle type in 3/44 (6.8%) and 3/44 (6.8%) of specimens, respectively (Table 1).
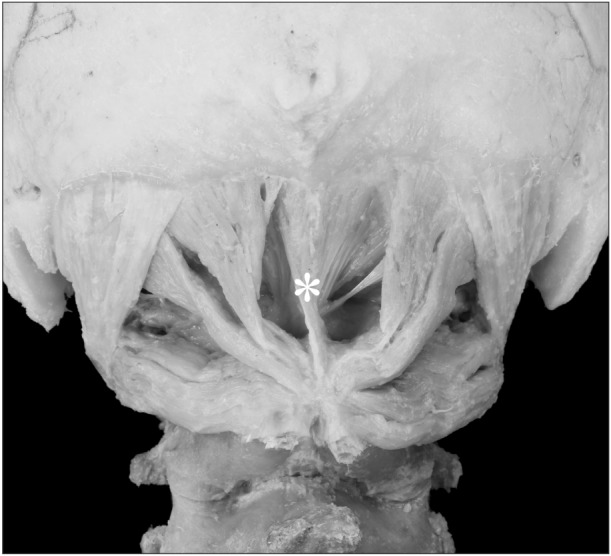
The case of RCPmi defect was bilateral (one cadaver; two samples). In one case, the anomalous muscle was located in the midline, where it originated from the spinous process of the second cervical vertebra and terminated on the inferior nuchal line of the occipital bone (Fig. 3). No anomalous muscle was seen in the OCS or OCI, which appeared as a typical muscle in all samples (Table 1). Features such as fat tissue, nerves, and blood vessels were seen between the muscle bellies of anomalous muscles.
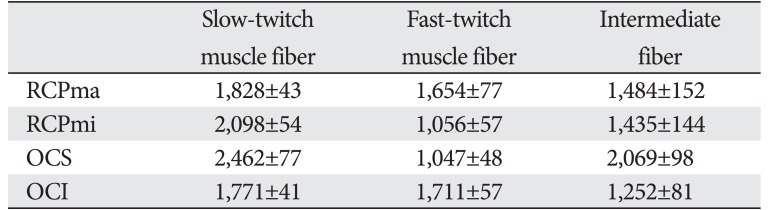
Mean CSA of muscle fibers
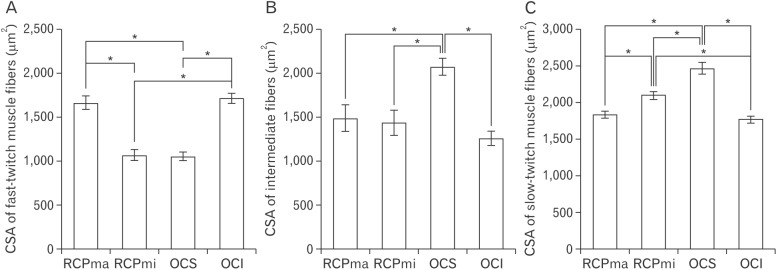
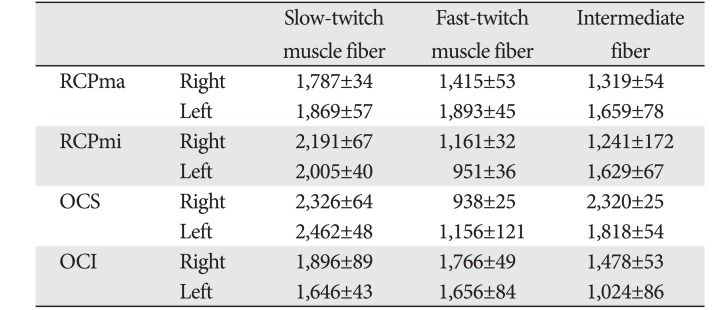
A comparison of the CSA of fast- and slow-twitch muscle fibers revealed that the CSA of the latter was at least two-fold greater than that of the former in RCPmi OCS (Table 2, Fig. 4). We were demonstrated no right-left difference (Table 3). The RCPma displayed a significantly greater CSA of fast-twitch muscle fibers than the RCPmi, and the OCI had a significantly greater CSA of fast-twitch muscle fibers than the RCPma. Meanwhile, the OCS had a significantly different CSA for intermediate fibers than the RCPma, RCPmi, and OCI. A comparison of the CSAs of slow-twitch muscle fibers revealed a significantly greater CSA in the OCS than in the other three muscles, and the RCPmi had a significantly greater CSA of slow-twitch muscle fibers than the RCPma and OCI (Fig. 5).
Discussion
The spine is generally surrounded by deep muscle and ligaments, but the atlanto-occipital and atlanto-axial joints contain no articular discs or interspinous ligaments [1516]. A likely reason for this is to enable highly flexible movement of the head instead of strong intervertebral immobilization. This suggests that the atlanto-occipital joint is controlled by the surrounding tissue to functionally maintain stability. The present study showed that slow-twitch muscle fibers have a CSA approximately greater than that of fast-twitch muscle fibers in all suboccipital muscles. Furthermore, these suboccipital muscles have a high density of muscle spindles, which, in addition to allowing flexible movement, act as specific sensory receptors [17]. The suboccipital muscles therefore act to functionally maintain the stability of the head while allowing delicate control of movement of the atlanto-occipital and atlanto-axial joints with a weak, sustained force.
Kamibayashi and Richmond [18] reported that the CSAs of the neck muscles were proportional to the height and weight of the individual. Furthermore, Ikai and Fukunaga [19] reported that muscle strength was proportional to the CSA of muscle fibers. Meanwhile, Rantanen et al. [20] demonstrated that no major age-related changes occur in the CSA of muscle fibers as a result of changes in the muscles of the erector spinae. The results for muscle fiber CSAs obtained in this study indicated that the RCPmi and OCS might provide more force than the RCPma and OCI, suggesting their potential roles as anti-gravity agonist muscles of the head.
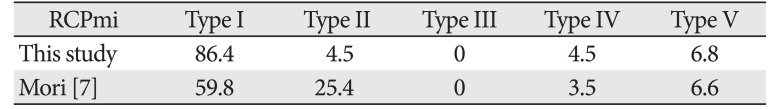
This study also revealed variations in the number of muscle bellies of the suboccipital muscles, which typically have only one muscle belly. Moreover, anomalous muscle was observed in 13.6% of RCPma and RCPmi samples, but was not observed at all in the OCS or OCI. Comparisons of the incidence of anomalous muscles in this study with that found by Mori [7] are presented in Tables 4 and 5. The presence of three muscle bellies was not described at all by Mori, but this feature was seen in 2.3% of the RCPma samples in our study, suggesting a large variation in the type of muscle anomaly. The incidence of two muscle bellies in the RCPmi in our study was about 21% lower, but the rate of muscle belly defects was about 1% higher. The incidence of typical muscle was also about 26% higher, revealing many cases of typical and missing muscle in the RCPmi. Katori et al. [21] reported that the mode of travel of structures such as nerves and blood vessels influences variations in the muscle belly. Because we observed features such as fat tissue, nerves and blood vessels between muscle bellies, the influence of embryological factors also seems likely.
Many previous studies have suggested that the suboccipital muscles are associated with chronic head and neck pain [2223]. Andary et al. [24] pointed out that atrophy of the suboccipital muscles is associated with chronic neck pain. Evaluation of the RCPmi by magnetic resonance imaging has shown that subjects with atrophied suboccipital muscles experience twice as much cervical vertebral dysfunction as control subjects [25]. Furthermore, in their clinical studies of chronic neck pain, Elliott et al. [262728] stated that fat tissue infiltrates the RCPmi as a result of damage to suboccipital muscle tissue. The pattern of the RCPmi defect in this study resembled that reported by Elliott et al. [262728]. Moreover, the RCPma was notably large in comparison with the other muscles in cases of type IV muscle defect, suggesting that the RCPma acted to compensate for RCPmi defects.
Mild neck pain can worsen when affected individuals have to work with visual display terminals (VDTs). Caution is therefore required with VDT work [29]. Individuals with anomalous suboccipital muscles could develop dysfunction if they are subjected to undue stress resulting from prolonged head extension.


 XML Download
XML Download