

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Fetal development of the external nose, ear, and eye involves a temporary plug formation resulting from the increased proliferation of keratinocytes, followed by canalization or recanalization mediated by a specific type of programmed cell death, i.e., cornification [12]. Recanalization of the anterior nose starts at 13 weeks and ends at 17 weeks of gestation [345]. Similarly, recanalization of the external auditory meatus starts at 13 weeks and continues until 16.5 weeks of gestation [67]. Likewise, the excretory tear duct of the eye, consisting of the lacrimal canaliculi, lacrimal sac and nasolacrimal duct, is formed by canalization of the cord-like structure of keratinized epithelial cells [89]. Canalization through a keratinized epidermal/epithelial tissue is also seen in the fetal perineal region such as the glandular urethra [1011]. In addition, although the embryonic stomodeum maintains its initial opening for the fetal mouth, a significant proliferation of oral and cutaneous keratinocytes is seen along the future lip margins and followed by the destruction to sculpture the upper and lower oral vestibules [3]. In those contexts, we should make clear the difference between the epidermal/epithelial plug and the usual cornification of skin. First, proliferation of keratinocytes continuously advances to fill the cavity or ductal lumen completely. Thus, the thickness of keratinocyte layer is much thicker than the usual skin. Second, according to Masumoto et al. [12], most or all of the keratinocytes most likely to carry cell nucleus in the plugs, in contrast to lost nuclei in the usual cornified layer of skin.
To our knowledge, the morphological difference between the fetal plug and usual skin was not examined immunohistochemistry. We hypothesized that, during a process of the plug formation, basal cells of the epidermis/epithelium may determine the final amount and extent of the keratinocyte mass according to the space of lumen. Therefore, in the present study, we paid attention to mesenchymal cells beneath the epidermis/epithelium. A change in intermediate filaments from cytokeratin (CK) to vimentin seems to be essential for epithelial-mesenchymal transition in normal embryonic development [1314] as well as cancer metastasis [1516]. In the present study, we used CK5/6, CK7, CK14, and CK19 for the epithelial marker: CK7 and CK19 are characterized by their positivity in the ductal epithelium and these are commonly used as a marker of tumors [1718]. CK5/6 and CK14 are not famous but they are considered to be kinds of maturation markers of the epithelial lining [1920]. All types of intermediate filaments seemed to be candidates to determine differentiation of the epidermis/epithelium: (1) vimentin is the most famous marker of mesenchymal tissues (reviewed by Galou et al. [21]); (2) glial fibrilary acidic protein (GFAP) and nestin are well-known markers of developing nerves (reviewed by Shibata et al. [22]). However, GFAP and nestin express also in mesenchymal tissues in and around fetal limb joints [23] and elastic cartilage tissues (reviewed by Katori et al. [24]). (3) Desmin plays a critical role in the initial attachment between striated muscle and tendons [25] and is thought to maintain the stability of mesenchymal cells in association with vimentin, another intermediate filament [26]. In addition, CD34 is a major marker of stromal (mesenchymal) and vascular progenitor cells in both adults and fetuses [2728]. However, the dermis and subcutaneous tissue also express CD34 [293031]. Consequently, we examined expressions of intermediate filaments and other mesenchymal markers beneath cornification in the fetal face.
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in 2013). Semiserial paraffin sections of the head (horizontal; 5 µm in thickness; 20–50 µm interval) were obtained from 5 mid-term human fetuses at 14–16 weeks (crown-rump length, 100–125 mm; 1 fetus at 14 weeks; 2 at 15 weeks; 2 at 16 weeks). These specimens were donated by their families to the Department of Anatomy, Yanbian University Medical College, Yanji, China, and their use for research was approved by the university ethics committee in Yanji (No. BS-13-35). These fetuses were obtained by induced abortion, after which the mother was orally informed by an obstetrician at the college teaching hospital of the possibility of donating the fetus for research; no attempt was made to actively encourage the donation. After the mother agreed, the fetus was assigned a specimen number and stored in 10% w/w neutral formalin solution for more than 1 month. Because of specimen number randomization, there was no possibility of contacting the family at a later date. After dividing the body into parts, head samples were decalcified by incubating at 4℃ in 0.5 mol/L ethylenediaminetetraacetic acid (pH 7.5) solution (Decalcifying Solution B, Wako, Tokyo, Japan) for 3–5 days, depending on the size of the sample.
From each of the 5 head specimens, we prepared 150–300 semiserial sections covering the entire face. One of every 5 sections were stained with hematoxylin and eosin to find sections including (1) the skin at the zygomatic or parotid region, (2) the external auditory meatus, (3) external nasal opening, (4) the lateral surface of the tongue, or (5) the putative lower lip or a lower margin of the mouth. After finding the sites, four of every five sections (i.e., unstained sections) were used for immunohistochemistry.
The primary antibodies used for immunohistochemistry were (1) mouse monoclonal anti-human cytokeratin (keratin) 5/6 or CK5/6 (M7237, 1:100, Dako, Glostrup, Denmark), (2) mouse monoclonal anti-human cytokeratin 7 or CK7 (1:100, N1626, Dako), (3) mouse monoclonal anti-human cytokeratin 14 or CK14 (1:50, LL002, Novo, Newcastle upon Tyne, UK), (4) mouse monoclonal anti-human cytokeratin 19 or CK19 (sc-6278, 1:100, Santa Cruz Biotechnology, Santa Cruz, CA, USA), (5) mouse monoclonal anti-human vimentin (1:10, M7020, Dako), (6) mouse monoclonal anti-human nestin (sc23927, 1:100, Santa Cruz Biotechnology), (7) rabbit polyclonal anti-human glial fibrilary acidic protein or GFAP (1:100, Z0334, Dako Cytomation, Kyoto, Japan), (8) mouse monoclonal anti-human desmin (1:100, M0760, Dako), (9) mouse monoclonal anti-human CD34 class II (M7165, 1:100, Dako), (10) mouse monoclonal anti-human proliferating cell nuclear antigen or PCNA (0879, Dako, 1:100), and (11) mouse monoclonal anti-human CD68 KP1 (M0814, 1:100, Dako). Autoclave pretreatment was not conducted because of the loose nature of the fetal tissues. The secondary antibody (Dako Chem Mate Envison Kit, Dako) was labeled with horseradish peroxidase (HRP), and antigen-antibody reactions were detected by the HRP-catalyzed reaction with diaminobenzidine. Counterstaining with hematoxylin was performed on the same samples. The negative control without a first antibody was set up for each of the specimens. Observations and taking photographs were usually performed with Nikon Eclipse 80 (Nikon, Tokyo, Japan).
Results
Observations of hematoxylin and eosin sections
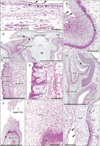
Fig. 1 displays topographical anatomy in and around the 5 sites examined. The zygomatic skin (Fig. 1A) was composed of (1) a thin but distinct, eosinophilic, homogeneous lamina or the stratum corneum and (2) the basal layer (stratum basale) and, between them, (3) the 2–3 layered keratinocytes as an intermediate layer. In the anterior nose (Fig. 1B, G), the lower lip (Fig. 1E, F) and the external auditory meatus (Fig. 1H, I), the intermediate, keratinocyte layer was very thick (more than 0.5 mm at maximum) with a tall basal cell layer. In spite of the thick keratinocyte layer, a surface homogeneous layer or the stratum corneum was thin or unclear in the nose, lip and ear. Notably, in these 3 sites, most of the keratinocytes carried cell nucleus.
In the external ear, the basal cell nuclei were large in the posterolateral aspect surrounded by elastic cartilages, but they were small in the anteromedial aspect continuous to the developing tympanic membrane. Thus, the latter calls appeared to be simple columnar (Fig. 1I). At the lip margin, the keratinocyte layer was very thick and connected with the contralateral side by a thin bridge (Fig. 1E). The lingual epithelium was characterized by abundant, deep protrusions of the basal layer into the internal, mesenchymal tissue (Fig. 1C, D). The epithelial protrusion was similar to the stratum spinosum of the adult skin because of the irregularly-shaped cells packed in a narrow area. No melanin granules were found in any cells at all sites examined.
Observations of immunohistochemistry for CKs
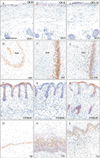
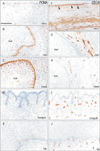
Basal cells were positive for CK19, CK14, and CK5/6 at the skin, external ear and nose, tongue and lip (Fig. 2). However, the positive cells for CK14 were restricted in the deep protrusion into mesenchymal tissues. Keratinocytes in the plug also usually expressed these 3 CKs, but in basal cells of the plug, positivity of CK19 was weaker than that of CK14 or CK5/6 (Fig. 2B, F, J). Expression of CK7 was seen in epithelia of the upper pharynx and auditory tube as well as the developing salivary glands (Fig. 3A–C). A stratum corneum covering the lingual surface also expressed CK7-positive (Fig. 3C).
Observations of immunohistochemistry for vimentin
In all sites examined, the vimentin-positive mesenchymal tissue was closely located to the basal layer of the epidermis or epithelium (Fig. 4). However, in the external auditory meatus and the lower lip margin, a thin homogeneous lamina, that was vimentin-negative, was evident beneath the basal layer of the epithelium (Fig. 4B, E). However, the lamina was thin and difficult to identify in the other 3 sites (Fig. 4A, C, D). Notably, in the external nasal opening and the external auditory meatus, vimentin-positive mesenchymal cells and their fragments were scattered both in the basal layer and the keratinocyte mass (Fig. 4B, C).
Observations of immunohistochemistry for nestin
Striated muscles were strongly positive near the zygomatic skin, the external nasal opening, the external auditory meatus and the lingual epithelium (Fig. 4F–I). Nestin-reactivity was also seen in mesenchymal cells and fibers especially near the basal layer: the reactivity was extremely strong around the external meatus as well as beneath the zygomatic skin. Notably, nestin-positive cells took a linear arrangement beneath the basal layer of the skin (Fig. 4F). In the external auditory meatus, a thin homogeneous lamina, that was nestin-negative, was seen beneath the basal layer (Fig. 4G).
Observations of immunohistochemistry for proliferating cell nuclear antigen or PCNA
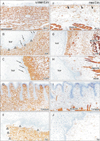
The basal layer of the zygomatic skin often or sometimes expressed PCNA: almost half of basal cells were weakly positive, while one of every 10 cells were strongly positive (Fig. 6A). The positive cells were likely to be adjacent in the basal layer. A thin, cornified layer of the skin contained a few positive cells. In the putative dermis and subcutaneous layer, the positive cells were scattered. Almost all basal cells at the external auditory meatus as well as all basal cells at the external nasal opening were strongly positive (Fig. 6B, C). Notably, in the plug, large keratinocytes in the basal two-third always contained PCNA-positive nucleus in the external nose and usually in the external ear (Fig. 6B, C). The deep protrusions of the basal layer of the lingual epithelium contained fewer positive cells than the primitive papilla (Fig. 6D). The lower lip margin contained several positive cells at the basal layer. Notably, near the basal layer of the lip, keratinocytes in the thick cornified layer carried PCNA-positive nucleus (Fig. 6E).
Observations of immunohistochemistry for CD34
Mesenchymal and/or fibrous tissues beneath the zygomatic skin and along the external auditory meatus expressed CD34 much more strongly than mesenchymal tissues in the nose, tongue and lip when they were compared in the same section (Fig. 6F–J). The zygomatic skin carried a CD34-positive cell monolayer beneath the basal layer: the lamina appeared to be composed of cells and fibers (Fig. 6F). The external auditory meatus also carried the similar lamina but it was thin and weakly positive (Fig. 6G). We did not find the lamina in the external nasal opening, the lingual epithelium and the lower lip margin. In these 3 sites, CD34-positive structures were limited to vessels (Fig. 6H–J). The vessels reached the basal layer in the lip margin, but most of them did not enter into the papilla.
Discussion
First, we summarized the present results. A stratum corneum was evident in the zygomatic skin and lingual surface, but the latter was positive for CK7. Between the stratum basale and stratum corneum of the primitive skin, there was a simple intermediate layer comprising of 2–3 layered, large keratinocytes. This basic configuration was common in the other 4 sites, but the intermediate, keratinocyte layer was much thicker than the zygomatic skin. A mesenchymal cell monolayer (nestin+, CD34+) was seen beneath the basal cells in the usual skin, but not in around the keratinocyte plug. Notably, in all 5 sites examined, all or most keratinocytes contained nucleus and, in the external ear and nose, they were positive for PCNA. Moreover, in the latter 2 sites, vimentinpositive mesenchymal cells appeared to migrate into the keratinocyte plug. The lingual epithelium was characterized by abundant, deep protrusions of the basal layer into the internal, mesenchymal tissue.
Keratin or CK is a well-known component of the epidermal and epithelial cells. However, to our knowledge, few study demonstrated the site-dependent difference in the fetal face: Shuler and Schwartz [32] carried the same question but antibodies were not defined at the date. In the present study, basal cells were always positive for CK19, 14, and 5/6 at any sites examined, but contrary to the skin, positivity of CK19 was weaker than that of CK14 or CK5/6 in the keratinocyte plug at the external ear and nose. The lingual epithelium was also different from the skin: the CK14-positive cells were restricted in the deep protrusion and the stratum corneum expressed CK7. The lingual epithelium was characterized by abundant, deep protrusions of the basal layer into the internal, mesenchymal tissues. The strange expression of CK7 seemed not to connect with the lingual epithelium-specific differentiation such as the late maturation of keratinocytes as well as the early development of taste buds [3334]. CK9 expression is known in thick and hard cornification such as seen at the foot-sole [35], but thick keratinocyte layers at the fetal face might be different in differentiation and development.
The most striking finding in the present study seemed to be vimentin-positive mesenchymal cells scattering in the keratinocyte plug. This morphology suggested a superficial migration of mesenchymal cells and, between the migrating cell and large keratinocyte, an epitheliummesenchyme interaction might be maintained in the plug for the continuous proliferation of keratinocytes. Conversely, when the keratinocytes fills the luminal spaces of the anterior nose and external auditory meatus completely, a pressure in the tissue was likely to increase to stop the superficial migration of vimentin-positive cells. A difference in the basal cell morphology was evident between the anterior and posterior aspects of the external auditory meatus: this fact might also suggest a contribution of the mesenchymal lining for determination of the epidermis. The tympanic cavity was adjacent to the anterior epidermis, while the posterior epidermis was surrounded by elastic cartilages. However, another interpretation seemed to be found in a difference in cell origin in external auditory epidermis [36]. In the lip margin, the amount of died and detached keratinocytes was likely to depend on the attaching area: the area seemed to be determined by the developing muscle mass in the lip. PCNA expression in the keratinocyte plug seems to be another striking feature. However, according to Scott et al. [37] and Onuma et al. [38], keratinocytes after final differentiation to cornification are likely to express PCNA. Therefore, in spite of the positivity of PCNA in the plug, some or most of the large keratinocytes were likely to lose proliferation activity. We needed to compare the results of PCNA staining with that of Ki67, but we had no residual unstained sections: this is a critical limitaion of this study.
No macrophages in the keratinocyte plug at the external ear and nose was one of the striking features. Because of no reaction of the keratinocyte plug found using deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end-labeling method, Masumoto et al. [12] hypothesized a substance-P mediated cell death, but they themselves found that non-specific immunoreactivity was likely to bring their result. Cornification is a highly specific type of programmed cell death [12]. The epidermal differentiation does not involve the pro-apoptotic caspases, but is associated with caspase-14 induction and processing [39]. Conversely, to understand the plug formation, a major point seemed not to be found in cell death with caspase but proliferation. Does an epithelial mesenchymal interaction provide a clue for the continuous proliferation of keratinocytes in the plug? The suggested superficial migration of mesenchymal cells into the plug (see the paragraph above) might be a way of resolution. Likewise, a mesenchymal cell monolayer (nestin+, CD34+) was absent around the keratinocyte plug. In contrast, a thin homogeneous, vimentin-negative lamina was evident beneath the basal layer at the external ear and lip margin. This difference seemed to be also important but, without further information of matrix in the lamina, our discussion could not expand.
CD34-positive fibrous tissues were evident beneath the skin and along the external auditory meatus in contrast to the anterior nose, tongue and lip those expressed CD34 only in the vascular wall. This regionalization of CD34-positve tissues were consistent with that described by Abe et al. [31] and Katori et al. [30]. However, a mesenchymal cell monolayer (nestin+, CD34+) characterized the usual skin. Overall, loss of a mesenchymal monolayer as well as superficial migration of mesenchymal cells might connect with the specific differentiation of keratinocytes in the plug. Finally, a further study seems to be necessary for tight junction proteins in and around the keratinocyte plug since a similar plug in squamous cell carcinoma provided an evidence [40].
 XML Download
XML Download