

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Anxiety, which may be understood as the pathological counterpart of normal fear, is manifested by disturbances of mood, as well as of thinking, behavior, and physiological activity [1]. The etiology of most anxiety disorders, although not fully understood, has come into sharper focus in the recent past. Serotonergic neurocircuitry is known to be involved in the development of anxiety [234]. 5-HT is believed to be involved in pathogenesis of anxiety, and plays the important role in mediating behavioral effects of anxiolytic drugs [567891011].
Serotonin type 2 (5-HT2) receptors have been implicated as possible targets to modulate anxiety behavior. Recently, numerous studies have revealed the involvement of 5-HT2A serotonergic receptor in the pathophysiology of anxiety [12]. Serotonin type 2A (5-HT2A) receptor knockout is reported to exhibit reduced anxiety responses, suggesting a role for the 5-HT2 receptors in establishing a baseline anxiety state [913]. The corticolimbic distribution of the 5-HT2A receptors might be critically involved in the neuropathology and treatment of a variety of psychiatric disorders including anxiety [79141516171819]. Various studies have indicated anxiolytic-like actions of 5-HT2 receptor agonists and anxiogenic-like effects of 5-HT2 receptor antagonists in diverse animal models [16202122232425].
5-HT2A receptor is located on both the excitatory and the inhibitory neurons in the rat hippocampus region, indicating that this receptor can participate in the information flow of the hippocampus region by acting on different neuronal populations [26]. The exact role of serotonin in the hippocampus region is largely unknown and functional studies concerning the effects of serotonin on the hippocampus region have been contradictory. One explanation for these contradictory results would be that serotonin has different effects on the hippocampus region depending on the serotonin receptor subtype involved [26]. However, it remains unknown whether 5-HT2 receptors contribute to the enhanced anxiety [27].
Steroid hormones have modulatory effects on the synthesis and release of several neurotransmitters like serotonin [2829] and affect serotonergic transmission in some extent by regulating the density of receptors in neuronal plasma membranes [303132]. Changing conditions such as steroid level may also exert different modulatory effects on the number and class of serotonin receptors present in the plasma membrane [33]. Androgens stimulate serotonin related gene expression [34] and they also participate in the modulation of function and activity in mature brain [35], as well as anxiety and depression [363738].
Steroid hormones like testosterone are able to influence the behaviour of people and animals [39]. In male rats, increased anxiety was observed in elevated plus maze and open field test after castration [40]. In contrast, after administration of testosterone to castrated rats, the anxiety was reduced [41]. Also, testosterone in the hippocampus of male rats is protective against orchidectomy-induced depressive-like behavior [42]. Testosterone plays profound roles in the regulation of the hippocampus structure and function in both rodents and human subjects [43]. For example, testosterone treatment significantly increased the spine synapse density in the hippocampus of rats [44] and regulated hippocampal spinogenesis [45]. Indeed, testosterone acts as a neurosteroid in neurons and can induce changes at the cellular level, which in turn leads to changes in behaviour, mood, and memory [46]. Testosterone also prevents neuronal death in different experimental models of neurodegeneration, and decreased testosterone levels in plasma may represent a risk factor for the development of neurodegenerative diseases in humans [474849].
Whereas the 5-HT2A receptor may be a potential therapeutic target for those disorders related to hippocampal dysfunction [50], the aim of the present study was to investigate the effects of chronic treatment of testosterone on anxiety and the number of 5-HT2A receptors–immunoreactive neurons in the hippocampus of gonadectomized male mice. We also examined the effects of testosterone on cell density (number of neurons and astrocytes) in the hippocampus of gonadectomized male mice.
Materials and Methods
Animals
Two-month-old adult male NMRI mice (n=36) were obtained from Pasteur Institute of Iran. The animals were kept in a controlled temperature room (22±3℃) with a 12:12 hour light/dark cycle (light beginning at 07:00–19:00 hour). They were given free access to rodent diet and water ad libitum and the number of animals in each cage was 4. Each animal was used only once. The experimental protocol was approved by the Ethic Committee of Golestan University of Medical Sciences, Iran.
Gonadectomy
Anesthesia was induced by intraperitoneal injection of 90 mg/kg ketamine and 10 mg/kg xylazine (Alfasan, Utrecht, Netherlands). After the onset of anesthesia, gonadectomy was performed through a ventral incision above the scrotum. Behavioral testing was initiated after a recovery period of 11 days.
Elevated plus maze test
The elevated plus maze (EPM) test was used for evaluating anxiety-related behavior in rodents [51]. The wooden EPM apparatus was consisted of opposite facing two open arms (50 cm×10 cm) and two closed arms (50 cm×10 cm×20 cm), connected by a central platform (10×10 cm). The whole maze was raised 50 cm above the ground level. Animals were tested in the maze in randomized order: the test was initiated by placing the mice on the central platform of the EPM apparatus facing towards one of the two open arms and allowed to explore it for 5 minutes. The number of entries into and time spent in each arm was recorded by an observer blind to the treatments. If mice placed all four paws on one of the four arms, an entry to the arms was registered. After 5 minutes test session for each mouse, the maze was wiped clean with 70% ethanol by damp cotton.
A number of classical parameters were collected during the session, as follows:
Open arm latency, that is, the time from placing the mice into the EPM apparatus until it entered one of the open arms
% Open arm time=Time spent in the open arms300
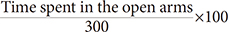 ×100
×100% Closed arm time=Time spent in the closed arms300
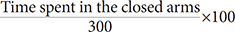 ×100
×100% Open arm entries=Number of open arm entriesNumber of total arm entries
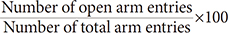 ×100
×100% Closed arm entries=Number of closed arm entriesNumber of total arm entries
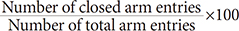 ×100
×100Total arm entries were measured as a relative pure index of locomotor activity.
Drug administration
Animals were randomly divided into six main groups: intact-no testosterone treatment (No T), gonadectomy (GDX)-No T, GDX-Vehicle, GDX-6.25 mg/kg testosterone (T), GDX-12.5 mg/kg T, and GDX-25 mg/kg T. Testosterone dissolved in DMSO (Merck, Darmstadt, Germany) and animals belonging to the GDX-Vehicle and GDX-testosterone treatment groups were received subcutaneous injections of vehicle (DMSO) or testosterone for 14 days. Testosterone doses were chosen based on previous studies [5253]. Mice subjected to EPM test at 30 minutes after last treatment.
Tissue preparation
After 14 days of treatment with testosterone (GDX-6.25 mg/kg T, GDX-12.5 mg/kg T, and GDX-25 mg/kg T groups) or DMSO (GDX-Vehicle group), and performing behavioral test, mice were decapitated and brain slices were prepared as described below. All mice were given a deep anesthesia with chloroform and transcardially perfused with 150 ml of saline followed by 600 ml of 4% paraformaldehyde solution. After perfusion, brains were removed carefully and fixed for at least 2 weeks in 4% paraformaldehyde. For tissue processing, brains were transferred to automate tissue processor apparatus (Did Sabz, Urmia, Iran). Following this session, samples of brain were embedded in paraffin and kept in refrigerator. Then, coronal serial 6-µm sections were obtained and the sections were divided into four sets for different staining separately.
Immunohistochemistry staining
The sections were stained with antibody against 5-HT2A serotonergic receptor, according to our previous report [54] with slight modifications. The sections were first incubated at 37℃ for 30 minutes, then, for deparaffinization, the sections embedded in xylene and hydrated with different degrees of ethanol. After these the samples were washed in purified water. The sections were then incubated with proteinase K solution (IHC World, Menlo Park, CA, USA) for 15 minutes at 37℃ in a humidified chamber and were allowed to cool for 10 minutes at room temperature. After several washes in washing buffer (phosphate buffered saline [PBS]/Tween 20 in 0.1% Triton X-100), the sections were blocked with peroxidase blocking solution (IHC World) for 10 minutes at room temperature and were then washed in washing buffer. Next, sections were blocked with avidin/biotin blocking solution (IHC World) for 30 minutes at room temperature and rinsed with PBS. Afterwards, sections were incubated with the 5HT2A receptor rabbit polyclonal antibody (1:500, Abcam Inc., Cambridge, MA, USA) for 70 minutes at 37℃. After several washes with washing buffer, the sections were incubated with secondary antibody (biotinylated goat anti-rabbit IgG antibody, Abcam Inc.) for 60 minutes at 37℃ and rinsed with washing buffer. Then, the sections were incubated with streptavidin HRP protein (1:5,000, Abcam Inc.) for 30 minutes at room temperature and rinsed with washing buffer again. The sections were then visualized using a DAB chromogen solution (IHC World) and counterstained lightly with Meyer's hematoxylin. Finally, the sections were dehydrated in ethanol, cleared using xylene and cover slip with entellan (Merck).
Phosphotangestic acid hematoxylin staining
Phosphotangestic acid hematoxylin (PTAH) staining was used for histological visualization of hippocampal astrocytes [555657]. The sections were first deparaffinized, rehydrated, and washed with distilled water. The sections were then incubated with Zenker's fixative for 1 hour at 50℃ and washed with tap water. Afterwards, the sections were placed in Lugol's iodine solution and then were bleached out iodine in 95% ethanol. The sections were then washed with distilled water. The sections were oxidized in 0.25% potassium permanganate and then were bleached in oxalic acid until white. Finally, the sections were placed in PTAH stain (Merck) for overnight at room temperature. After staining, slides were dehydrated quickly through two changes each of 96%, 100% ethanol, and cleared using two changes of xylene and cover slip with entellan. In this method, the astrocytes appear blue-violet and the neurons become pink.
Cresyl violet staining
The sections were embedded in xylene for deparaffinization and hydrated through a graded series of ethanol and finally rinsed with distilled water. Next, the sections were stained in 0.02% Cresyl violet (Sigma, St. Louis, MO, USA) solution for 5 minutes and then rinsed quickly in distilled water. Finally, the sections were dehydrated in 96%, 100% ethanol for 2–3 minutes, cleared in xylene for 3–5 minutes and cover slip with entellan [55].
Hematoxylin and eosin staining
The sections were stained with hematoxylin and eosin staining for dark neurons in accordance with routine laboratory procedures. Dark neurons were diagnosed by the neuronal shrinkage, cytoplasmic eosinophilia, nuclear pyknosis, and surrounding spongiosis [58].
Image processing and cell counting
All images from hippocampal areas including CA1, CA3, and DG areas were recorded by using a digital camera (DP 72, Olympus, Tokyo, Japan) attached to the light microscope (BX51, Olympus). Using ImageJ software, the appropriate grids were superimposed on the pictures. To measure astrocytes and dark neurons, a field of 30,000 µm2 was counted manually in CA1, CA3, and DG areas of the hippocampal formation. Also, the neural cells labeled for 5-HT2A receptor and neurons were counted manually in 30,000 µm2 for CA1 and CA3 areas and also 4,800 µm2 for DG area [59].
Results
The effects of gonadectomy and testosterone therapy on anxiety in plus maze test
As seen in Fig. 1, gonadectomy affected male mice anxiety-like behavior in the elevated plus maze. One-way ANOVA revealed that gonadectomy in GDX-No T group decreased % open arm time (Fig. 1A), % open arm entries (Fig. 1C), and the pure index of locomotor activity (Fig. 1F) in comparison to Intact-No T group. It was also found that gonadectomy in GDX-No T group increased % closed arm time (P<0.05) (Fig. 1B) and open arm latency (Fig. 1E) in comparison to Intact-No T group. No significant change in % closed arm entries (Fig. 1D) was observed following castration. These data showed that gonadectomy induced an anxiogenic-like effect in male mice.
It was also demonstrated that subcutaneous injections of testosterone at dose of 6.25 mg/kg/day after gonadectomy increased % open arm time, % open arm entries and the pure index of locomotor activity (Fig. 1A, B, F) while it decreased significantly the % closed arm time (P<0.05) (Fig. 1B), and open arm latency (Fig. 1E), compared to the GDX-No T group. But, this dose of testosterone did not alter % closed arm entries (Fig. 1D). This indicates that low dose of testosterone partially reverses gonadectomy induced anxiogenic-like effects.
Data analysis showed that administration of intermediate dose of testosterone (12.5 mg/kg/day) after gonadectomy increased significantly % open arm time (P<0.01) (Fig. 1A), % open arm entries (P<0.01) (Fig. 1C), and the pure index of locomotor activity (P<0.01) (Fig. 1F). Also, this dose of testosterone decreased significantly % closed arm time (P<0.01) (Fig. 1B), % closed arm entries (P<0.01) (Fig. 1D), and open arm latency (P<0.05) (Fig. 1E), compared to the GDX-No T group. This suggests that the intermediate dose of testosterone can induce an anxiolytic-like effect.
Moreover, the results demonstrated that treatment with high dose of testosterone (25 mg/kg/day) after gonadectomy is unable to cause a significant change in % open arm time (Fig. 1A), % open and closed arm entries (Fig. 1C, D), open arm latency (Fig. 1E), and the pure index of locomotor activity (Fig. 1F), compared to the GDX-No T group. Furthermore, this dose of testosterone slightly decreased % closed arm time (P<0.05) (Fig. 1B) compared to the GDX-No T group. It can be concluded that high dose of testosterone may induce an anxiogenic-like effect.
Effect of testosterone on density of 5-HT2A receptor-immunoreactive neurons
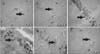
Fig. 2 shows coronal sections of mice hippocampus (CA1 area) stained by immunohistochemistry with an antibody against a 5-HT2A serotonin receptor.
Gonadectomy decreased the density of 5-HT2A receptor-immunoreactive neurons in the hippocampal CA1, CA3, and DG areas compared to Intact-No T mice (P<0.001) (Table 1). Treatment with different doses of testosterone for 14 days increased density of 5-HT2A receptors–immunoreactive neurons in all hippocampal areas of GDX-mice. These increases in density of 5-HT2A receptor-immunoreactive neurons were very low at GDX-6.25 mg/kg T group and it had not significant difference with GDX-No T mice. In the hippocampal CA1, CA3, and DG areas, the post hoc LSD test revealed that the density of 5-HT2A receptor-immunoreactive neurons were significantly higher in GDX-12.5 mg/kg T and GDX-25 mg/kg T groups compared to GDX-No T mice (P<0.001) (Table 1). The high density of 5-HT2A receptor-immunoreactive neurons in hippocampal CA1, CA3, and DG areas observed by administration of 25 mg/kg dose of testosterone.
Astrocytes density changes in CA1, CA3, and DG areas of the hippocampus
We used PTAH staining to determine the hippocampal astrocytes (Fig. 3). The results showed that the density of the astrocytes in the GDX-No T and GDX-Vehicle groups were reduced by the gonadectomy at CA1, CA3, and DG areas of the hippocampus, compared with the Intact-No T group (Fig. 4), indicating that gonadectomy decreases the density of hippocampal astrocytes.
Moreover, low dose of testosterone caused a significant increase in the density of astrocytes at the CA1 (P<0.001) (Fig. 4) and CA3 (P<0.01) (Fig. 4) regions of the hippocampus when compared to the GDX-No T group.
The density of astrocytes increased significantly in the CA1 and CA3 areas of the hippocampus in comparison to the GDX-No T group after treatment with intermediate dose of testosterone (P<0.001 and P<0.01, respectively, at 12.5 mg/kg/day) (Fig. 4).
Subcutaneous injections of testosterone (25 mg/kg/day) after gonadectomy resulted in a significant increase in density of astrocytes in CA1 and CA3 areas of the hippocampus when compared to the GDX-No T group (P<0.001 and P<0.05, respectively) (Fig. 4).
In the DG region, the density of astrocytes was also increased in all testosterone-treated groups when compared with the GDX-No T group, but this difference was statistically significant after treatment with high dose of testosterone (at 25 mg/kg/day) (Fig. 4).
The data analysis also revealed that gonadectomy decreased the density of astrocytes and testosterone treatment with different doses increased their density.
Neurons density changes in CA1, CA3, and DG areas of the hippocampus
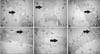
Fig. 5 shows hippocampal neurons in the CA1 area stained with cresyl violet staining. As we showed in Table 2, gonadectomy decreased the density of pyramidal neurons in CA1 area, while partially decreasing the density of these cells in the CA3 region. It also caused a significant increase in the density of granular cells in the DG area compared to the Intact-No T and the GDX-No T group (P<0.05) (Table 2).
Post-gonadectomy testosterone therapy with low does increased significantly the density of pyramidal neurons at CA1 and CA3 areas, compared to the GDX-No T group (P<0.05 and P<0.01, respectively, at 6.25 mg/kg/day) (Table 2).
Our findings also demonstrated that subcutaneous injections of testosterone at a dose of 12.5 mg/kg/day increased significantly the density of pyramidal neurons in CA1 compared to the GDX-No T group (P<0.01, at 12.5 mg/kg/day) (Table 2). Moreover, intermediate dose of testosterone slightly increased the density of these cells in the CA3 area (Table 2).
The high dose testosterone treatment caused notable increase in the density of pyramidal neurons in the CA1 area in comparison to the GDX-No T group (P<0.05, at 25 mg/kg/day) (Table 2), although it did not cause any significant changes in the CA3 area (Table 2).
Administration of testosterone with different doses decreased the density of granular cells in the DG area, compared to the GDX-No T group. Further analysis also revealed that maximum significant decrease observed at the low dose of testosterone-treated group (P<0.05, at 6.25 mg/kg/day) (Table 2).
Furthermore, it was revealed that treatment of the GDX male mice with low and intermediate doses of testosterone may prevent neuron reduction, while administration of high dose testosterone cannot prevent this decline in the GDX male mice.
Effect of testosterone on density of dark neurons
Fig. 6 shows dark neurons in the hippocampal CA1 area stained with H&E staining. The production of dark neurons was studied in the hippocampal CA1 and CA3 areas as well as in the DG represented in Fig. 7. Histological studies indicated a significant increase in the mean density of dark neurons in two hippocampal area, including CA1 and CA3 in all testosterone treated animals compared to GDX-No T animals. The mean density of dark neurons in the hippocampal CA1 area was significantly higher in GDX-6.25 mg/kg T and GDX-12.5 mg/kg T groups (P<0.05) and GDX-25 mg/kg T group (P<0.001) compare to GDX-No T animals. In addition, the mean density of dark neurons in the hippocampal CA3 area was significantly higher in GDX-12.5 mg/kg T animals in comparison to GDX-No T group (P<0.001). However, testosterone-treated groups in the DG region showed no significant changes compared to GDX-No T animals.
Discussion
Gonadectomy induced anxiety-like behavior in adult male mice. Current results demonstrated diverse effects on anxiety after fourteen days treatment with testosterone in a dose dependent manner. Administration of intermediate dose of testosterone (in GDX-12.5 mg/kg T group) was associated significantly with anxiety-like behavior. There were no significant changes in behavioral test after application of both lower and higher doses of testosterone (in GDX-6.25 mg/kg T and GDX-25 mg/kg T groups) in bilaterally castrated mice.
Consistent with these data, McDermott et al. [60] showed that castration increases anxiety only if the mice were castrated after puberty. Furthermore, gonadectomy results in increased anxiety-like behavior [406162].
Previous studies have shown that testosterone is associated with anti-anxiety behavior, and case studies indicated that testosterone substitution in hypogonadism reduces anxiety [63]. Moreover, at intact adult male Wistar rats, exogenous testosterone (5 mg/kg, intramuscularly) has anxiolytic effects, at least as assessed in the light-dark transition task, 24 hours after testosterone injection [64]. Also, most studies demonstrated that administration of testosterone or its metabolites was anxiolytic in aged intact mice [65], GDX rats [4052616667], and intact male rats [66].
In another study, repeated treatment with testosterone propionate (0.5 and 1 mg/rat, subcutaneous, 4 injections), produced an anxiolytic-like effect in a probe-burying test [41]. Also, in the present study, we showed that injection of testosterone with the dose of 12.5 mg/kg for 14 days caused anxiolytic behavior.
Roohbakhsh et al. [53] demonstrated testosterone increased % open arm entries at the dose of 10 mg/kg (subcutaneous injection) in male rats. Also in another study, testosterone with a dose of 25 mg/pellet (60-day slow release testosterone) in isolated GDX male and ovariectomized female rats had no effect on the number of entries into the open arms [52]. Our results demonstrated that chronic treatment with testosterone, with a dose of 12.5 mg/kg increased significantly % open arm entries; hence testosterone at this dose has an anxiolytic-like effect in GDX male mice.
Our findings indicated that administration of testosterone at doses of 6.25 mg/kg and 12.5 mg/kg increased pure index of locomotor activity. However, recently, it has been shown that injection of testosterone with doses of 5, 10, and 20 mg/kg (subcutaneous injection) produced no significant change in the locomotor activity [53].
There was a relationship between behavioural and histological data in our study which indicates that the most increase in the number of astrocytes in mice hippocampus was seen in 6.25 and 12.5 mg/kg/day doses of testosterone compared to the other doses. These results are similar to Emamian et al.'s study [57], which represented that learning impairment caused by intra-CA1 microinjection of testosterone (80 µg/0.5 µl) increased the number of astrocytes in the CA1 region of the rat hippocampus.
Similarly, Del Cerro et al. [68] demonstrated that castration of adult male rats resulted in a significant decrease in the extension of glial fibrillary acidic protein (GFAP)–immunoreactive processes and in contrast, treatment with testosterone enhanced the extension of these processes. Our findings are also consistent with previous studies showing that testosterone can affect astrocytes in the hippocampus. For example, testosterone administration to newborn females and castration of newborn males showed significant alteration in GFAP expression in the hippocampus and hypothalamus [6970].
Our results also showed that treatment with different doses of testosterone for 2 weeks, compensated gonadectomy-induced reduction of astrocytes quantities in the GDX-No T group and increased their numbers in all testosterone-treated groups. Although, in another study by Barreto-Estrada et al. [71], it was found that both early and delayed administration of testosterone (5 mg/kg, subcutaneous) resulted in a significant decrease in the number of vimentin-immunoreactive astrocytes and volume fraction of major histocompatibility complex-II immunoreactive microglia after brain injury in male rats [71].
Some recent reports have suggested a relationship between neurogenesis and anxiety-related behaviour [72737475]. A couple of these studies demonstrated that deficit of hippocampal neurogenesis leads to increased anxiety-related behaviours [727374]. However, the molecular mechanisms that link hippocampal neurogenesis to anxiety disorder remain poorly understood [72].
Castration of male mice had no effect on cell proliferation, but it did cause a decrease in the number of immature neurons in the DG area [7677]. However, castration with chronic mild unpredictable stress may reduce hippocampal cell proliferation [78]. Other studies showed that castration with chronic mild unpredictable stress [77] and/or castration in aggregation with social isolation stress [79] reduced neurogenesis among adult male rats. In accordance to these results, we also found that the number of neurons can be decreased in the CA1 area after gonadectomy, while it can stimulate neurogenesis in the DG area of GDX male mice.
In agreement with our data, Allen et al. [80] showed that gonadectomy increases neurogenesis in male adolescent rhesus macaque hippocampus. They concluded that the absence of adolescent testicular hormones had no effect on cell proliferation, but increased cell survival by 65% and indices of immature neuronal differentiation by 56% in GDX monkeys compared to intact monkeys.
The discrepancies in the effects of castration on cell proliferation in the hippocampus between different studies could be due to the time between castration and perfusion and/or the length of hormone withdrawal [78].
The actions of gonadal hormones on hippocampal cell proliferation and neurogenesis are quite complex [81] and some studies support the conclusion that testosterone does not necessarily enhance adult neurogenesis [79]. Several studies demonstrated that testosterone implants given to intact male rats for 21 days had no effect on hippocampal neurogenesis [3782], suggesting that testosterone may differentially modulate neurogenesis at adolescence [83]. Another study showed that testosterone represents as important mediators of cell proliferation in the subventricular zone of adult rats [84]. Similar to these reports, our results demonstrated that testosterone has a suppressive effect on neurogenesis in the DG area of mice hippocampus.
It has been previously described that steroid hormone could play dramatic role in inhibition of neuronal death through reduction in stress level. Our data from behavioral test showed that the animals after gonadectomy have higher levels of stress. Although in treated animal who received testosterone, lower level of stress has been found. In GDX-12.5 mg/kg T group we observed a reduction in stress-like behaviors. As we also observed significant changes in the number of 5-HT2A receptor-immunoreactive neurons, therefore it could be concluded that serotonin receptors (5-HT2A) may play a significant role in stressful behaviors.
The results of immunohistochemistry demonstrated that castration of male mice reduced the number of 5-HT2A serotonergic receptor-immunoreactive neurons in the CA1, CA3, and DG regions of the hippocampus. Chronic treatment with testosterone also increases the number of 5-HT2A serotonergic receptor-immunoreactive neurons in the hippocampus in all testosterone-treated groups. It seems that these increases at the 5-HT2A receptor-immunoreactive neurons are related to the dose of testosterone.
In agreement with our results, other studies demonstrated that gonadectomy in female and male rats decreased, while testosterone or estrogen increased 5-HT2A receptor mRNA levels in frontal cortex [85] and dorsal raphe nucleus [8687]. Gonadectomized female and male rats demonstrated reduced density of 5-HT2A receptor binding sites and or 5-HT2A receptor density in frontal, cingulate and primary olfactory cortex and nucleus accumbens and this could be increased by testosterone or estrogen treatment [85868788].
An increase of 5-HT2A receptors was found in cortical brain regions following administration of estrogen alone in human studies [89] or combined estrogen and progesterone [9091]. Furthermore, in another study, estradiol treatment in combination with progesterone (for 2 weeks) increased 5-HT2A receptor mRNA expression in the CA1 and CA2 region of the ventral hippocampus at the ovariectomized adult female rats [92].
According to our finding, testosterone can plays an essential role in increasing the number of 5-HT2A serotonergic receptor-immunoreactive neurons, this effect of testosterone can be implicitly explained by the actions of its more physiologically active metabolites, dihydrotestosterone and estradiol. Indeed, testosterone is converted to estrogen by the enzyme aromatase. It can thus modulate the serotonergic system by binding to estrogen receptors. In addition, the estrogen receptor β is expressed on serotonergic neurons that are therefore targets of testosterone converted to estrogen by aromatase (e.g., 5-HT2A receptor) [9394]. As well as, throughout the hippocampus of rodents express aromatase in a steroid- and sex-independent manner. Given that the hippocampal formation is rich in both androgen and estrogen receptors, it is possible that testosterone may exert anxiolytic effects via both androgen-dependent and estrogen-dependent mechanisms within this brain region [42].
Vermeire et al. [11] exhibited lower 5-HT2A receptor binding index in the frontal and temporocortical regions (both left and right) and right occipital cortex in the dogs with anxiety disorders compared with the reference dogs. The reason for the lower binding index may be that the receptor is down regulated because of serotonergic activation in anxiety disorders in dogs. Finally, on the basis of these results, it is not possible to conclude whether the lower density of 5-HT2A receptors is causal to, or a consequence of, anxiety disorders in dogs [11].
In fact, some studies have shown the involvement the 5-HT2 receptors in anxiety [95]. Masse et al. [24] and de Paula Soares et al. [25] have reported that the stimulation of the 5-HT2A/2C receptor in specific brain regions causes anxiolytic-like effect in animal models.
Global disruption of cortical 5-HT2A receptor signaling in mice reduced inhibition in conflict anxiety paradigms without affecting fear-conditioned and depression-related behaviors [9]. Zhang et al. [96] demonstrated that 5-HT2A mRNA was decreased by gonadectomy in ventromedial hypothalamic nuclei, with reversal of the decrease again seen following testosterone replacement (150 µg/kg/day, intraperitoneal, 7 days).
The changes of other subtypes of 5-HT receptor induced by steroids in the hippocampus of ovariectomized rats were studied. Immunodetection of these receptors were indicated that exposure to estradiol (50 µg/kg, subcutaneous) and the combination of estradiol and progesterone significantly reduced the 5-HT5A–like immunosignal in the CA1 region of hippocampus while progesterone (7.5 mg/kg, subcutaneous) did not induce changes. Also, exposure to the combination of estradiol and progesterone or estradiol alone increased the 5-HT2C immunosignal in the CA1 area of hippocampus [33]. Furthermore, our findings demonstrated that chronic treatment of testosterone increased the number of 5-HT2A receptors–immunoreactive neurons in hippocampus.
Our behavioral results suggested that the density of 5-HT2A receptor-immunoreactive neurons may play a crucial role in the induction of anxiety-like behavior. As reduction in such receptor expression have shown to significantly enhanced anxiety behaviors. However, replacement of testosterone dose dependently enhances the number of 5-HT2A receptors–immunoreactive neurons and interestingly also reduced anxiety like behaviors.







 XML Download
XML Download