

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
During dentinogenesis, cranial neural crest cell–derived odontoblasts play a role in the secretion of pre-dentin and dentin following terminal differentiation [1]. Dentin is a major component of tooth and is produced by odontoblasts. During the late bell stage of tooth development, odontoblasts differentiate from ectomesenchymal cells of the dental papilla under the influence of the inner dental epithelium [2]. Moreover, odontoblasts then produce extracellular matrix, including collagen type I as well as non-collagenous proteins such as dentin sialophosphoprotein (Dspp), osteocalcin (OC), and dentin matrix protein 1 (Dmp1) which facilitate the mineralization of the dentin matrix [34].
Transforming growth factor-beta (TGF-β) superfamily, insulin-like growth factors, WNTs, fibroblast growth factors, and other kinds of growth factors are involved in differentiation of dental papilla cells into odontoblasts during tooth development [5]. TGF-β is a multifunctional regulator of a variety of cellular functions, including cell proliferation, differentiation, apoptosis and matrix synthesis [6]. TGF-β/bone morphogenetic protein (BMP) signaling has been implicated to have specific roles during tooth development as seen various disturbances with its disruption by tissue-specific targeting of odontoblast and dentin formation. In a previous study, Wnt1-Cre;Tgfbr2 mice have shown delayed odontoblast differentiation and decreased dentin thickness [7]. Moreover, targeted inactivation of Bmp2 or Bmp4 also showed phenotypes of impaired odontoblast maturation and tooth root defects [89]. These data suggested that TGF-β/BMP signaling plays critical role in odontoblast differentiation and dentin formation.
TGF-β superfamily consists of TGF-βs, BMPs, activins, and other related proteins, and TGF-β/BMP signaling transduction pathway maintains cell proliferation, differentiation, apoptosis, migration, and reconstruction of proteins [10]. Smad4, a key mediator of TGF-β/BMP signaling, functions as a multifunctional regulator for cranial neural crest cell migration, proliferation, differentiation, protein reconstruction, and immune response, or other physiological functions [1011] and expresses in oral epithelium and dental mesenchyme during tooth development [12]. To study the functions of Smad4 during full tooth development, tissue-specific gene targeting technology like conditional knockout strategy is essentially required to overcome disadvantage of Smad4-null mice, which is arrested at E7.5–E8.5 with early stage of tooth development [13].
Here, we generated and analyzed the tissue-specific conditional Smad4 disruption mice under the control of Osr2 and OC promoter to investigate the role of TGF-β/BMP signaling in odontoblast differentiation and dentin formation.
Materials and Methods
Mouse strains and tissue preparation
All experimental procedures were approved by the animal Welfare Committee of Chonbuk National University. Smad4-floxed allele (Smad4fl/fl), Osr2Ires-Cre, and OC-Cre mice have been previously described [141516]. Tissue specific activities of Osr2Ires-Cre and OC-Cre have been reported in dental mesenchyme and odontoblasts [1516]. Rosa26 (R26R) reporter mice [17] were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). To generate Osr2Ires-Cre:Smad4fl/fl (Osr2CreSmad4) and OC-Cre:Smad4fl/fl (OCCreSmad4) mice, Osr2Ires-Cre:Smad4fl/+ and OC-Cre:Smad4fl/+ (control) mice were crossed with Smad4fl/fl mice, respectively. Genotyping of mice was carried out by allele-specific polymerase chain reaction as previously described using the following oligonucleotide primers: Smad4 floxed (Smad4a, 5'-AAG AGC CAC AGG TCA AGC AG-3'; Smad4b, 5'-GGG CAG CGT AGC ATA TAA GA-3'; Smad4c, 5'-GAC CCA AAC GTC ACC TTC AC-3'), OC-Cre (Cre1, 5'-ATC CGA AAA GAA AAC GTT GA-3'; Cre2, 5'-ATC CAG GTT ACG GAT ATA GT-3'). Osr2Ires-Cre (Osr2Ires-Cre1, 5'-GAA TTC GCC AAT GAC AAG ACG CTG-3'; Osr2Ires-Cre2, 5'-CTA CAA GGA TCT AGC ACA TGC TG-3'), Rosa26R (R1295, 5'-GCG AAG AGT TTG TCC TCA ACC-3'; R523, 5'-GGA GCG GGA GAA ATG GAT-3'; R26F2, 5'-AAA GTC GCT CTG AGT TGT TAT-3'). To analyze the level of Cre activity, Osr2Ires-Cre, OC-Cre mice were crossed with Rosa26R mice, and the mandibles (P0 and P8) of the double-transgenic mice were processed for X-gal staining, as described previously [18].
Tissue preparation and histology
For histology analysis, the mice at the age of P0 to P28 were sacrificed and their heads and mandibles were carefully dissected. Tissues were fixed in 4% paraformaldehyde and decalcified in 10% ethylenediaminetetraacetic acid/phosphate buffered saline solution for 1 to 4 weeks at 4℃. The decalcified tissues were dehydrated through a graded ethanol series, embedded in paraffin, and sectioned at 5 µm thickness. Slides were stained with hematoxylin and eosin.
Immunohistochemistry
For immunohistochemistry, sections were treated with 3% hydrogen peroxide and incubated with rabbit polyclonal antibodies against osterix (Osx; 1:200, Abcam Inc., Cambridge, MA, USA), Dspp (1:200, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), phosphate regulating endopeptidase homology on the X chromosome (Phex; 1:50, Sigma-Aldrich, St. Louis, MO, USA), Dmp1 (1:750, Takara Bio Inc., Shiga, Japan), biglycan (Bgn; 1:800, Dr. Larry Fisher). Histostain Plus primary kit (Zymed Laboratories, San Francisco, CA, USA) was used according to the manufacturer's instructions.
Kidney capsule transplantation
The tooth germ of the mandibular at E14.5 was dissected from embryo. The explants were placed on filters supported by metal grid in a tissue culture dish and cultured for 1 day during genotyping. The host mouse was anesthetized using pentobarbital (0.5 mg/10 g body weight) and the explants were grafted under the kidney capsule according to standard procedure. Two weeks after transplantation, the host mice were sacrificed and the grafts were processed for histological analysis.
Results
Localization of Cre recombinase activity for gene targeting during tooth development
The Osr2Ires-Cre and OC-Cre transgene directed their specific activity of Cre recombinase at dental mesenchyme and odontoblasts respectively, as β-galactosidase activity was observed in the dental mesenchyme of Osr2CreR26R double transgenic mouse at E14.5 (Fig. 1A) and in odontoblasts of mandibular molars in OCCreR26R double transgenic mouse at P8 (Fig. 1B, C). Based on these results, we crossed Osr2Ires-Cre transgenic mice with Smad4fl/fl mice for targeting Smad4 ablation at the stage of initial coronal dentin formation. OC-Cre transgenic mice were used for targeting the stage of initial root dentin formation and long term observation of coronal dentin formation.
Impaired odontoblast differentiation with Smad4 disruption in dental mesenchyme
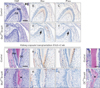
Since odontoblast differentiation was impaired following the disruption of Smad4 in dental mesenchyme, histological differences in odontoblast were compared with the mandibular molar of Osr2CreSmad4 and control mice (Fig. 2A, D). At newborn stage, Osr2CreSmad4 mice have a layer of non-polarized cuboidal cells with centrally located nucleus during coronal dentin formation while control mice have polarized odontoblasts. In addition, dentin matrix was absent in Osr2CreSmad4 mice (Fig. 2D). Immunohistochemistry of mandibular molars revealed that lower or no Osx and Phex were expressed in the odontoblasts of Osr2CreSmad4 mice (Fig. 2E, F) while highly expressed in differentiating odontoblast and odontoblastic processes in control mice (Fig. 2B, C). Osr2CreSmad4 mice die within a day after birth, precluding investigation of tooth development at later stage. To determine whether the abnormal odontoblast differentiation in Osr2CreSmad4 is due to delayed development, we performed kidney capsule transplantation. Mandibular first molars of embryo at E14.5 were collected and cultured for one day. By following the genotype results, it was transplanted on the kidney capsule of wild type mice and observed after 2 weeks (Fig. 2G–L). Interestingly, the tooth development of Osr2CreSmad4 mice was delayed even with normal enamel formation with well differentiated ameloblasts. However, enamel and dentin were formed with well differentiated ameloblasts and odontoblasts in control mice. Interestingly, ectopic hard tissue with cellular inclusion was formed by odontoblasts with a distinct difference from former produced dentin Osr2CreSmad4 mice (Fig. 2G, J). Cellular inclusion was commonly detected in ectopic hard tissue at cervical region of root dentin in the OCCreSmad4 mice targeting at the stage of initial root dentin formation (Fig. 2M, N). Immunohistochemistry after kidney capsule transplantation revealed that Osx was highly expressed in differentiating polarized odontoblasts in control mice while expressed in matrix forming cells and the cells trapped in dentin matrix in Osr2CreSmad4 mice (Fig. 2H, K). However, the expression of Phex was obviously decreased in the odontoblast of Osr2CreSmad4 mice when compared to control mice after kidney capsule transplantation (Fig. 2I, L). These results indicated that inactivation of TGF-β/BMP signaling in odontoblasts may disturb the differentiation processes of odontoblasts during tooth development.
Targeted ablation of Smad4 in odontoblasts leads a disturbance of dentin formation
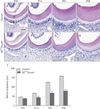
Ablation of Smad4 in odontoblasts was established in OCCreSmad4 mice as seen the specific activity of Cre activity in odontoblasts of mandibular molars by OCCreR26R double transgenic mouse (Fig. 1). Histological analysis of mandibular molars revealed that dentin thickness was definitely decreased in OCCreSmad4 mice compared to control mice. In addition, the odontoblasts have shorter height and flattened morphology in OCCreSmad4 mice than in control mice, implying the loss of odontoblast polarity by ablation of Smad4 in odontoblasts. At the age of P10, coronal dentin thickness did not show any remarkable difference between control and OCCreSmad4 mice (Fig. 3A, E, I). Interestingly, coronal dentin thickness was slightly increased in the OCCreSmad4 mice while dramatically increased in control mice at the age of P14, P21, and P28 (Fig. 3B–D, F–H). These results indicate that inactivation of TGF-β/BMP signaling in odontoblasts may disturb dentin matrix production during dentin formation.
Molecular changes in odontoblasts and dentin matrix with disruption of Smad4
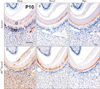
To determine molecular changes in odontoblasts and dentin formation following ablation of Smad4, immunohistochemical staining was performed with mandibular tissue sections of control and OCCreSmad4 mice at P10. Dspp was slightly expressed in odontoblasts and dentinal tubules in OCCreSmad4 mice while highly expressed in control mice (Fig. 4A, D). Dmp1 was observed in dentin matrix of OCCreSmad4 mice with strong expression while weak expression in control mice (Fig. 4B, E). Bgn, a proteoglycan, exhibited specific expression at pre-dentin in both control and OCCreSmad4 mice (Fig. 4C, F).
Discussion
In this study, we investigated the significance of Smad4, a key intracellular signaling mediator of TGF-β/BMP signaling, by tissue-specific ablation under the control of Osr2 or OC promoter during tooth development. We found that during dentinogenesis the ablation of Smad4 with two types of gene targeting by different Cre activation commonly results in disturbed differentiation of odontoblasts and abnormal bone-like dentin formation, which observed at the stage of initial coronal dentin formation as well as initial root dentin formation.
TGF-β superfamily is comprised of many members, such as TGF-βs, Nodal, Activin, and BMPs. TGF-β/BMP signaling transmits initial signals across the plasma membrane through the formation of heteromeric complexes of specific type I and type II serine/threonine kinase receptors. The type I receptor is phosphorylated and then activates specific type II receptor. Activated type I receptors initiate intracellular signaling through phosphorylation of specific Smad proteins, R-Smads. Activated R-Smads form a complex with Co-Smad and Smad4 and then translocate into the nucleus to direct transcriptional response [19]. TGF-β/BMP signaling plays an important role in regulating extensive processes including cell proliferation, differentiation, apoptosis, migration, and extracellular matrix remodeling [620]. From long ago, TGF-β/BMP signaling has been implicated to have an important role during dentin formation as seen that cranial neural crest cell–derived odontoblasts secret pre-dentin and dentin following terminal differentiation [21]. Odontoblasts are differentiated from the dental mesenchymal cells in dental papilla under the influence of the inner dental epithelium. Odontoblasts become tall and in columnar shape with differentiation [21]. Smad4, the intracellular mediator for the TGF-β/BMP signaling, plays important role in regulating early tooth development [22]. Among the TGF-β superfamily, TGFβ-1, TGFβ-2, TGFβ-3, BMP2, BMP4, BMP7, and follistatin are expressed in the inner enamel epithelium, dental papilla and in polarizing and functional odontoblasts. Exogenous TGFβ-1, BMP2, BMP4, and BMP7 can induce odontoblast differentiation and dentin formation in dental papilla cells in vitro [23242526]. These data indicate that TGF-β/BMP signaling play critical roles in odontoblast differentiation and dentin formation. In our data, Osr2CreSmad4 mice exhibited disturbed differentiation of odontoblasts and abnormal bone-like structures instead of dentin formation. The results suggest that conditional inactivation of Smad4 cause morphological and functional deficiency in odontoblasts during odontoblast differentiation and dentin formation, implying that Smad4-dependent TGF-β/BMP signaling plays a significant role in odontoblast differentiation and dentin formation during tooth development. In addition, impaired dentin formation was commonly exhibited in different types of Smad4 ablation models such as Dspp-Cre;Smad4, OC-Cre;Smad4, and Col-Cre;Smad4 at the crown, cervix, and root furcation area of molars respectively [27]. These abnormalities suggest the requirement of Smad4-dependent TGF-β/BMP signaling for appropriate dentin formation. Also, exogenous TGF-β1 regulates Dspp and Dmp1 expression in odontoblast cell lines [2829], indicating that TGF-β/BMP signaling regulates dentin matrix secretion. In our results, OCCreSmad4 mice exhibited thinner crown dentin and bone-like root dentin structures in the cervical region when compared to control mice. This abnormality suggests the failure of appropriate odontoblast differentiation and resultant reduction of dentin matrix apposition.
Taken together, our results suggest that Smad4-dependent TGF-β/BMP signaling plays a critical role in odontoblast differentiation and dentin formation during tooth development.

 XML Download
XML Download