

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Recently our group demonstrated a significantly greater proportion of tyrosine hydroxylase (TH)–positive, sympathetic nerve fibers in the frontal and auriculotemporal nerves than the other trigeminal nerve branches in elderly donated cadavers [1]. Therefore, parts of the human face around mouth and nose are unlikely to be innervated by sympathetic nerve fibers passing through nerve trunks but by artery-bounded nerve fibers. However, we have no information about composite fibers of cutaneous branches of the nasociliary nerve. During the proximal course in the orbit, the nasociliary nerve is richly communicated with the ciliary ganglion as well as the short ciliary nerves [2345]. Moreover, the human ciliary ganglion contains both TH-positive neurons and neuronal nitric oxide synthase (nNOS)-positive neurons [6]. Therefore, being different from the frontal nerve, the nasociliary nerve is likely to contain both sympathetic and parasympathetic nerve fibers.
A change in the human forehead skin circulation is known in response to arousal emotion to increase sympathetic nerve activation [78]. The forehead is innervated by the frontal and nasociliary nerves: both are branches of the first division of the trigeminal nerve. Near the forehead, smooth muscles, and glands in the eyelid also requires parasympathetic nerve supply, but details of the nerve course were not yet described [9]. Consequently, the aim of this study was to immunohistochemically identify sympathetic and parasympathetic nerve fibers in skin nerves of the first division of the trigeminal nerve.
Materials and Methods
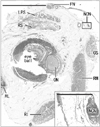
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000) and it was approved by the Human Research Ethical Committee by the dental school. We dissected 20 cadavers donated for the annual student dissection courses at Tokyo Dental College (15 males, 5 females; 71–98 years old; mean age, 85 years old at death). The cause of death in each case had been ischemic heart failure. The donated cadavers had been fixed by intravenous injection of non-neutralized 10% (v/v) formalin solution and preserved in 50% ethanol solution for more than 3 months. From each of the cadavers, we obtained an entire right orbital content including the eyelids and a part of the forehead skin and, after routine procedures for paraffin-embedded histology, we first prepared 5-µm-thick frontal section including posterior half of the eyeball and stained with hematoxylin and eosin (Fig. 1). After identification of the nasociliary nerve as well as the frontal nerve in the posterior site, we further prepared multiple sets of sections for immunohistochemistry. Sections were provides at 4–5 sites at 1–2 mm interval toward the eye lids and forehead. For identification of the sectional planes, one of the set always included subcutaneous tissues around the trochlea of the superior obliquus muscle. In addition, at the superolateral wall of the ethmoidal sinus, we prepared a tip of the anterior ethmoidal nerve and artery from each of the cadavers.
According to Hinata et al. [10], the primary antibodies used for nerve immunohistochemistry were (1) rabbit polyclonal anti-human nNOS (1:200, Cell Signaling Technology, Beverly, MA, USA), (2) mouse monoclonal anti-human vasoactive intestinal polypeptide or vasoactive intestinal polypeptide (VIP; 1:100, sc25347, Santa Cruz Biotechnology, Santa Cruz, CA, USA), and (3) rabbit polyclonal anti-human TH (1:500, ab152, Millipore-Chemicon, Temecula, CA, USA). We tried to perform these three types of immunohistochemistry using adjacent sections, but sometimes we used near sections (not adjacent) due to a failure of the sectioning and/or immunostaining. Antigen retrieval was performed using microwave treatment (500 W, 15 minutes, pH 6). The secondary antibody (incubation for 30 minutes; 1:1,000, Histofine Simple Stain Max-PO, Nichirei, Tokyo, Japan) was labeled with horseradish peroxidase (HRP), and antigen-antibody reactions were detected by the HRP-catalyzed reaction with diaminobenzidine. Counterstaining with hematoxylin was performed on the same samples. The negative control without a first antibody was set up for each of the specimens. The short ciliary nerves in the sections were used for the positive control. Observations and taking photographs were usually performed with Nikon Eclipse 80 (Nikon, Tokyo, Japan), but photos at the ultralow magnification (less than ×1 at the objective lens) were taken using a high grade flat scanner with translucent illumination (Epson scanner GTX970; Epson, Tokyo, Japan).
Results
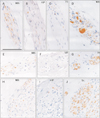
Identification of the nasociliary nerve was not difficult because, at the peripheral course, it ran near the superior obliquus muscle with a concomitant artery. After finding the nerve in a posterior section including the posterior pole of the eye ball (Fig. 1), we traced it toward the eyelid and forehead skin using semiserial sections. We included the trochlea of the superior obliquus muscle into one of the sections to make sure of the sectional planes. Immunohistochemistry of nNOS was successfully conducted in 12 of 20 cadavers, but the reactivity was not seen in short ciliary nerves (positive control) in the other 8 cadavers. The results shown below will be based on the 12 cadavers with successful results.
In the available 12 specimens, throughout the peripheral course from a level of the posterior pole of the eyeball, via a site near the trochlea and toward the skin, the nasociliary nerve consistently contained abundant nNOS-positve fibers. However, VIP-positive fibers were always few in number (Figs. 2, 3). TH-positive fibers considerably varied in number between specimens. We did not find a difference in the nNOS immunoreactivity between sites before and after an origin of the anterior ethmoidal nerve from the nasociliary nerve. Actually, we did not find nNOS reactivity in a fragment including the anterior ethmoidal nerve and artery (data not shown). The frontal nerve always contained abundant TH-positive fiber, but no or few VIP- and nNOS-positive fibers were included. The short ciliary nerves contained all three types of fibers. Although we found individual variations in TH-positive fiber contents, there appeared not to be the correlation with age or gender. Because of the longitudinal or oblique sections of nerve fibers, the morphometrical analysis was difficult.
Discussion
Many research groups had paid attention to a critical role of nitric oxide synthase to control blood supply in the skin [11121314]. However, an immunohistochemical demonstration of nNOS along the cutaneous nerve course seemed to be limited to Ibba-Manneschi et al. [15]. Using frozen sections of skin specimens from systemic sclerosis patients and healthy volunteers, they displayed a lower magnification views of subcutaneous nerves running toward the dermis. However, in the figures, we were not able to identify the positive nerve fibers or the other associated cells. Schulze et al. [16] demonstrated a co-existence of NOS and VIP in cutaneous nerve fibers in human face and forearm, but their results were not based on immunohistochemistry but enzyme histochemistry of NOS. Therefore, the present study might be the first demonstration of nNOS-positive nerve fibers in and along a skin nerve course.
A significant difference in composite nerve fibers between the frontal and nasociliary nerves seemed to be explained by a fact that the latter nerve is most likely to supply nNOS-positive fibers to smooth muscles and glands in the eyelid and upper nasal epithelium. However, the nNOS-positive fibers were also likely to play a role in emotional sweating in the forehead [78]. In contrast to sympathetic nerve fibers to skin [1718], it was difficult to find a report of nNOS expression in skin nerves. We had also conducted many immunohistochemical studies of the human perineum, but we did not find such fibers in skin nerves from the pudendal nerve [1019]. Likewise, in contrast to TH-positive fibers to skeletal muscles [20], Hosaka et al. [21] did not find nNOS-positive fibers in muscle nerves in the human body.
One reason of no or few reports on the parasympathetic peripheral nerve course was found in a difficulty in nNOS immunostaining using specimens from formalin-treated donated cadavers after long preservation [22]. Another reason seemed to exist in the so-called cholinergic sympathetic innervation [23]. Cholinergic parasympathetic fibers can express kinds of sympathetic nerve markers such as neuropeptide Y and/or dopamine-beta-hydroxylase [24]. However, according to our long trials, the cadaveric specimens after long preservation are not available for immunostainings of choline acetyltransferease, neuropeptide Y, and dopaminebeta-hydroxylase. This is a critical study limitation of this study. A study using biopsy specimens from volunteers could bring further information of parasympathetic nerve course to the skin. A new technique such as signal amplification systems [25] may be helpful.
 XML Download
XML Download