

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The penile and scrotal raphe is a subcutaneous fibrous plate extending from the penile frenulum, along the penile shaft and scrotum and toward the anus. In mice, the urorectal septum of endodermal origin plays a role of zipper to close the bilateral genital folds [12]. Thus, in mice, the midline seam or raphe is originated from the urorectal septum. However, since physiological hypospadias is evident in human fetuses, a contribution of the urorectal septum is still unclear in the penile raphe. An epithelial-mesenchymal interaction has been hypothesized for the delayed fusion under control of androgen [34]. In contrast, in the human perineum between the scrotum and anus, the urorectal septum is most likely to provide the perineal raphe as in mice. In human mid-term fetuses, a future scrotum was difficult to identify because of no descent of testis [5]. Thus, in the present study, we will apply a term "perineal raphe" to a raphe behind the root of penis.
Early development of the anus from the cloaca has been a major subject in embryology [67891011]. However, to our knowledge, only one researcher [12] described details of fetal development of the raphe possibly due to early stages observed by the other researchers. According to him, the human raphe appears at 9 weeks of gestation and becomes thicker and continuous with the anus at 12–13 weeks. Notably, van der Putte [12] did not consider the raphe as a remnant of fusion of the bilateral genital folds but a secondary formation after disappearance of the latter. Therefore, the first aim of this study was to describe fetal development of the male perineal raphe behind the root of penis.
In adults and children, the penile and scrotal raphe vary in shape and thicknmess between individuals and the anomalies include pearly, pigmented, prominent, or wide raphe [13]. The anomalies sometimes include a "skin groove" extending from the hypospadic urethra to the anus [1415]. Likewise, the perineal groove is known as a rare female anomaly consisting of a wet skin grove from the vestibule to the anus. This anomaly has been considered to result from (1) a failure of the genital folds to fuse, (2) a relic of the open cloaca, and (3) a failure of the urorectal septum to develop downward [1617]. In this context, a midline fusion of the bilateral genital folds seems to be hypothesized in the female perineum. However, the female genital folds are not able to fuse together and, instead, they are widely separated by the vestibule in female fetuses [18]. Surprisingly, in both genders, a thick perineal raphe or its variation is often evident in combination with various types of anorectal malformations [19]. Therefore, the second aim of this study was to examine whether the raphelike structure was present or absent behind the fetal vestibule. In the mid-term human fetus, the vagina does not descend to reach the vestibule [20].
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). We observed serial paraffin sections (5–10µm in thickness) of 30 fetuses (17 males and 13 females; crown-rump length 48–115 mm or approximately 10–15 weeks): they were composed of 9 males and 4 females at 10 weeks, 4 males and 5 females at 12–13 weeks, and 4 males and 4 females at 14–15 weeks. The sectional planes were frontal (20 specimens) or sagittal (10 specimens). Since these sagittal sections were slightly tilted to the frontal planes, midline structures such as the symphysis pubis and perineal raphe were able to be identified as linear structures. The sections had been stained with hematoxylin and eosin, Masson trichrome or azan as a part of the Medical Museum Collection, Georg- August-Universität Göttingen (4 specimens, all sagittal) and Institute of Embryology, Universidad Complutense Madrid (the other 26 specimens).
For observations of the Blechschmidt collection in the Medical Museum of Georg-August-Universität Göttingen [21], we did not need approval by the ethics committee of the institute. Our use of sections in the collection kept at the Institute of Embryology, Universidad Complutense Madrid [22] was approved by the university ethics committee (No. B08/374). Observations and taking photographs were performed with Nikon Eclipse 80 (Nikon, Tokyo, Japan) in the museum or institute.
Results
Observations of male specimens
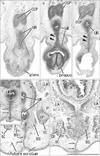
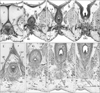
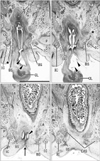
At 10 weeks, the perineal raphe was present in the superficial or posterior side of the proximal urethra. Notably, the initial raphe was continuous with and took a direction same as the inferior or posterior wall of the urethra after closure of physiological hypospadias (Fig. 1). In the collection in Madrid, we did not find a cross section of the penile urethra. Thus, we were not able to demonstrate clearly an absence of the penile raphe at the early stage. However, the penile urethra appeared to be as short as the initial perineal raphe (Fig. 1A vs. Fig. 1D). Depending on the tilting of sectional plane, the future scrotum was sometimes identified as a thick subcutaneous tissue in the inferior side of the pubic ramus (Fig. 1E). Since the thickened tissue was often difficult to identify because of no descent of testis (Fig. 2), instead of the term "scrotum," we will use "the root of penis" in the observations below. Near the anus, a midline septum was sandwiched by the bilateral corpus spongiosum penis as well as by the bilateral bulbospongiosus muscles (Fig. 2A–D).
At 12–13 weeks, the raphe became distinct but wavy in the subcutaneous tissue at and behind the root of penis (Fig. 2E–H). The bulbospongiosus muscle increased in mass and extended laterally toward the ischiocavernosus muscle and posteriorly toward the external anal sphincter (EAS) (Fig. 2F, G). However, in those specimens at 10–13 weeks, the EAS was still thin and small and identified as scattering muscle fibers in the subcutaneous tissue (Fig. 2D, G). At 14–15 weeks, the perineal raphe as well as its deep continuation (i.e., the intermuscular septum) was thick and almost straight, depending on the extent of growth of the bulbospongiosus muscles (Fig. 3). The posterior parts of the muscle were attached to and communicated with the EAS (Figs. 3C, 4). The perineal raphe ended at the anterior margin of the EAS (Figs. 3D, 4E). The urethral groove of the penis was completely closed until 12 weeks (i.e., closure of physiological hypospadias). In addition, throughout the ages examined, an intermuscular space was present between the bulbospongiosus and ischiocavernosus muscles (Figs. 2A, E, 3B, 4E).
Observations of female specimens
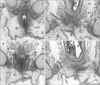
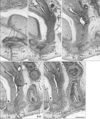
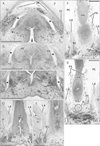
In all female specimens examined, the vaginal descent along the urogenital sinus or urethra was not completed: the inferior end of the vagina was still above the level of the inferior margin of the pubic symphysis. At 10–13 weeks, the large phallus or initial clitoris protruded anteriorly, making it difficult to distinguish from a penis with physiological hypospadias (Fig. 5). The inferior end of the vagina or the sinovaginal bulb was identified as a solid tissue mass on the posterosuperior side of the vestibule (Figs. 5A, 6, 7). In 2 specimens, we found a short epithelial fusion or seam at the posterior end of the vestibule (Fig. 5C), but this midline fusion did not connect with the bulbospongiosus muscle. At 14–15 weeks, the muscle elongated superiorly and posteriorly along the vestibular bulb (Fig. 6).
In all specimens, the bilateral bulbospongiosus muscles were separated widely by the vestibule (Figs. 5D, 6B, C), except for the anterosuperior end near the crus clitoris. The muscle did not attach to the EAS, but there was a loose tissue mass interposed in front of the EAS (Fig. 7). As described in males, an intermuscular space was consistently seen between the bulbospongiosus and ischiocavernosus muscles (Figs. 5A, 6C). Overall, there was no evidence of a raphe-like structure in the fetal female perineum. The gender difference in topographical anatomy of the bulbospongiosus muscle and raphe is schematically represented in Fig. 8. Finally, we did not find a candidate of the perineal body in the present fetuses.
Discussion
In the present study, we incidentally found a short epithelial fusion or seam at the posterior end of the vestibule. However, in front of the EAS, a large midline mesenchymal tissue did not contain a raphe-like structure. In the male perineum, the raphe grew as a superficial extension of the midline or intermuscular septum between the bilateral bulbospongiosus muscles. The raphe always reached the EAS. Conversely, according to Arakawa et al. [23], the subcutaneous part of the EAS tends to develop near and along the perineal raphe in male fetuses [23]. Therefore, an abnormal raphe was likely combined with abnormal perineal muscles including the EAS. In the female perineum, the bilateral muscles were separated widely by the vestibule. Without reinforcement by the midline muscular septum, the female raphe could not maintain its growth even if the seed appeared at the posterior end of the vestibule. This situation was somewhat similar to a difficulty in fetal development of the female circular rhabdosphincter [1820]. Due to a long distance to the bulbospongiosus muscles, a major driving force for growth of the penile raphe might be an increased thickness of the septum between the bilateral corpus cavernosum penis.
In relation to fetal development of perineal midline structures in females, we introduced an anomalous perineal groove (see the "Introduction"). In female fetuses, a large mesenchymal tissue from the urorectal septum existed in front of the EAS and it appeared not to be involved in the growing vestibule. Therefore, rather than a failure of the genital folds to fuse, a failure of the urorectal septum to develop downward seemed to cause the abnormality in the female perineum. Because it is often lined by rectal columnar epithelium, not by the squamous epithelium [16], the perineal groove seemed not to be a relic of the open cloacal duct. The developing anal sinus is likely to contribute to provide the columnar epithelium of the groove since a long sinus sometimes extends anteriorly to reach the smooth muscle wall [24]. In addition, a candidate of the perineal body was not found in the present fetuses since it is originated from the longitudinal smooth muscle layer of the rectum at 20 weeks or later [2325].
A great limitation of the present study was found in lack of suitable transverse sections of the penis at 10–12 weeks. Using such sections, a further study was necessary to ensure no penile raphe immediately after the closure of physiological hypospadias. However, because of sections with a routine staining, it seemed to be difficult to deny a possibility that, after epithelial-mesenchymal transition, the epithelial seam for closure differentiates into the initial penile raphe.

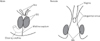
 XML Download
XML Download