

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Advances in brain imaging have provided new insights into the relationship between motor learning and brain structure. Imaging investigations have revealed that motor learning and training can evoke plastic changes in brain structures that are associated with distinct training demands, and such learning-induced structural alterations have been demonstrated to arise in both the gray matter (GM) and white matter (WM) in the human brain.
Longitudinal investigations focused on motor learning have revealed structural plasticity in specific WM regions. For example, Juggling training increased fractional anisotropy (FA), a measure of the directionality of water diffusion to measure the WM microstructure, in the WM underlying the intraparietal sulcus [1]. Furthermore, bilateral upper extremity motor skill training increased FA in the posterior and anterior limbs of the internal capsule, the corona radiata, and the body of the corpus callosum [2], and complex visuomotor rotation training was found to result in increase in FA below the primary motor cortex [3]. Conversely, learning of a complex whole-body balance task decreased FA in prefrontal WM regions [4].
In our previous study, we found that elite basketball athletes showed larger volumes of vermian lobule VI-VII (declive, folium, and tuber) in the cerebellum after long-term intensive training. This finding indicated the important role of the cerebellum in motor learning and coordination [5]. However, we did not determine whether GM or WM contributed to the increased volume of the cerebellar vermis.
In this study, we further examined whether the increased volume of this region induced by extensive motor learning in elite basketball athletes is due to a larger GM or WM volume. The results of this study are expected to lead to a better understanding of learning-induced structural plasticity in the human brain. In addition, this study design is advantageous compared to pooling large datasets from elite athletes.
Materials and Methods
Subjects
Nineteen male elite basketball athletes group (BG) involved in national and international competitions were recruited from several university teams in South Korea through direct visiting interviews. Members of the BG with heights over 190 cm were excluded from this investigation to reduce the height variation among subjects. Twenty healthy male subjects who did not engage in any sports-related activity were selected as a control group (CG) through advertisements on Korea University web pages and in the local community newspapers.
Before magnetic resonance imaging (MRI) scanning, we obtained information about alcohol consumption and the type of alcohol consumed, as well as the use of nicotine and other drugs through a questionnaire. Neurological examination was performed by a neurologist at Korea University Medical Center, and any subjects with potential neurological impairments were excluded from the study. All subjects had healthy general medical and neurological profiles, and none of the subjects presented postural and vision problems.
The duration of training and practice time per day were checked in the BG, and body heights and weights were measured in all subjects. Table 1 presents the average age, height, and weight of all right-handed subjects. Written informed consent was acquired from all the subjects, and all procedures were approved by the Institutional Review Board of Korea University.
MRI acquisition
MRI was executed on a 1.5-tesla Magnetom vision instrument (Siemens, Erlangen, Germany) using T1-weighted magnetization prepared rapid acquisition gradient echo sequence (repetition time, 9.7 milliseconds; echo time, 4 milliseconds; flip angle, 12 degrees; slice thickness, continuous 2.0 mm; field of view, 250 mm; number of slices, 90; and matrix, 256×256). After the acquisition of magnetic resonance (MR) images, the Digital Imaging and Communications in Medicine (DICOM) files were transferred to an IBM compatible PC.
Volumetry
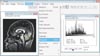
The region of interest (ROI) areas of the cerebellar vermis was manually delineated on two sagittal slices of MR images (the mid-sagittal slice and the parasagittal slice in which the cerebral aqueduct became continuous with the fourth ventricle) using the ImageJ program (National Institutes of Health, Bethesda, MD, USA) to measure the volumes of the ROI's for GM and WM (Fig. 1). The numbers of pixels of marked region of each ROI were calculated by histogram analysis and then copied to an Excel sheet to distinguish between GM and WM based on gray-scale density (Fig. 1). A pixel density value from 26 to 100 was regarded as GM, and values greater than 105 were regarded as WM. The GM and WM volumes for all ROIs were calculated by multiplying the total numbers of pixels of each ROI by the voxel size (0.89×0.89×2.00 mm).
Vermian V1 included lobules I-V (the lingual, centralis, and culmen), vermian V2 included lobules VI and VII (the declive, folium, and tuber), and vermian V3 comprised lobules VIII-X (the pyramis, uvula, and nodulus). All ROIs were independently drawn for each subject by two trained raters that were strictly blinded to the subjects' identity and assignment, and were checked by a neuroradiologist. Inter-rater reliability was tested using two-tailed Pearson correlation. Inter-rater correlation coefficients were over 0.90 for all ROIs. We normalized these ROIs by calculating the relative GM and WM ROI volumes for each subject as a percentage of their intracranial volume in order to minimize inter-subject variability.
Statistical analysis
Statistical analyses were performed with the SPSS software package version 12.0 (SPSS Inc., Chicago, IL, USA). The Student's t test was performed to determine the effect of athletic proficiency on the absolute and relative GM and WM volumes of the ROIs. All analyses were two-tailed, and a P<0.05 was considered as statistically significant. All data are represented as mean±SD.
Results
Comparison of the absolute volumes of each ROI between the BG and CG showed no significant differences (Table 2). Comparison of the relative volumes of each ROI between the BG and CG showed significant differences in the relative WM volumes of V2 (Table 3). The relative GM volume of the V1, relative GM volume of the V2, relative GM volume of the V3, relative WM volume of the V1, and relative WM volume of the V3 showed no group differences.
Discussion
In the present study, we identified larger WM volumes of the vermian lobules VI and VII (the declive, folium, and tuber) in the cerebellums of elite basketball athletes compared to those of controls. We suggest that the larger volume of vermian lobule VI-VII observed in elite basketball athletes in our previous study [5] was driven mainly by the WM of this region. The results of our study suggest that the structural alterations of WM in this region appear to have been induced by long-term extensive motor learning, and that the WM of this region might play an essential role in coordination.
Elite athletes usually begin sports training in early childhood, and tend to practice several hours per day intensively. Therefore, highly trained elite athletes could provide a good model to investigate training-induced structural plasticity in the human brain. A number of studies using MRI have revealed structural differences in the GM and WM of multiple brain regions in response to a practiced task between elite athletes and control subjects, which might reflect the effects of long-term motor learning [678910]. However, inconsistent results have been reported for both GM and WM structures.
In GM structures, elite judo players had a larger GM volume in the frontal lobe which is related to motor planning and execution, as well as in regions of the prefrontal cortex, which is related to working memory and cognitive processes [6]. In addition, professional and low-handicap golfers had larger GM volumes in the fronto-parietal network, including premotor and parietal areas [7], and badminton athletes showed a greater GM concentration in the right anterior and posterior lobes of the cerebellum [8]. However, decreased GM volumes were observed in the left premotor cortex, supplementary motor area, putamen, and the superior frontal gyrus of professional female ballet dancers [9]. With respect to WM structure, world-class gymnasts showed higher FA values in the bilateral corticospinal track [10]. By contrast, professional and low-handicap golfers showed a lower WM volume and FA values in the vicinity of the corticospinal tract at the level of the internal and external capsule and in the parietal operculum [7], and professional ballet dancers showed a smaller WM volume in the corticospinal tracts, internal capsules, corpus callosum, and left anterior cingulum and lower FA values in the left and right premotor cortex [9].
Some longitudinal studies have shown a cause-and-effect relationship with respect to motor learning such as juggling training, bilateral upper extremity training, visuomotor rotation training, and balance training, and their direct influence on WM structure in the adult human brain [1234]. Moreover, inter-individual variation in the WM microstructure is related to performance variation in a bimanual coordination task [11]. The cerebellum plays an important role in motor learning [1213], and motor learning-induced structural plasticity in the WM of the cerebellum has also been reported. MRI analyses of adult Japanese macaque monkeys showed an increased WM volume in the cerebellum after tool-use training [14]. Pianists and string players, as examples of individuals with specific motor expertise, showed a plastic change of WM in the cerebellum [15]. Indeed, FA in the WM of the cerebellum positively correlates with learning scores of a visuo-motor task [16]. Motor coordination was shown to be involved in individual differences in WM integrity in the cerebellum in karate black belts groups [17]. These findings indicate that structural properties of WM in the cerebellum are relevant to motor learning.
WM, which lies under the GM cortex, consists of millions of axons that link neurons in different regions into functional circuits. The white color originates from the myelin that covers the axons. Myelin is crucial for the conduction of electrical impulses, and its deficit can damage neuronal transmissions [18]. Changes in WM, including the number of axons, diameter of axons, myelin thickness, axon branching, axon trajectories, and myelination, influence the speed of impulse propagation [19]. Thus, improving the velocity or synchrony of impulse conduction could be an essential feature of motor learning, and could in turn influence the improvement of performance with motor learning [1920].
Previous studies revealed that experience or training could change the WM structure. An enriched environment increases the number of unmyelinated and myelinated axons, as well as oligodendrocytes that form myelin in rats [212223242526], and the size of the corpus callosum in rhesus monkeys [27]. Environmental effects on the WM structure have also been demonstrated in the human brain. Early experience increases the WM structure in the internal capsule and frontal lobes in newborn infants [28]. Tool use training induces extension of the axonal branches in the cerebral cortex of monkeys [29], and the amount of electrical activity within an axon could regulate its degree of myelination [3031].
In this study we attempted to match the age, height, and weight as much as possible because these variables could affect the cerebellar volume. There was significant difference in weight between BG and CG. Cerebellar volume significantly affected by body height [32]. Therefore, it is not likely that the weight variable affected the result of this study.
The results of our study, combined with the previous demonstration of WM alterations in the cerebellum of musicians [15] and tool-use trained monkeys [14], suggest that extensive motor learning may induce morphological alterations in the WM of the cerebellum. Basketball involves complex motor skills, which require bimanual and eye-hand coordination for dribbling and shooting a ball. Thus, participation in a basketball competition at a high level of competition requires very high motor proficiency. These skills are central to basketball achievement and are therefore extensively and specifically trained. It seems that elite basketball athletes have highly developed motor skills, and such long-term extensive motor learning would evoke structural plasticity in the WM of vermian lobules VI-VII in the cerebellum. Such WM plasticity in this region of athletes may be crucial to support the increased functional demands for athletic achievement. Extensive motor learning may induce neural activity, which in turn may increase the number of axons, axon diameter, thickness of myelin, axon branching, axon trajectories, and myelination in this region. These microscopic changes might be reflected in the WM of this region in elite basketball athletes to improve the speed or synchrony of impulse transmission to support increased motor demands, including coordination and balance control [1920].
However, T1-weighted MRI has relatively low sensitivity, because MRI cannot reflect the structures of WM in the human brain as diffusion tensor imaging. Therefore, the findings of WM volume differences must be interpreted with caution. In addition, it is technically difficult to apply our MR images acquired from non-isometric voxels to a voxel-based morphometry technique based on statistical parametric mapping. Further studies are required to compare imaging data with the post-mortem histological measures that underlie these macroscopic finding in elite athletes, in order to more clearly establish the anatomical basis of these motor learning-induced morphological alterations in the human brain. In addition, the connectivity of the cerebellum with the red nucleus and cerebral cortex also requires further study to clarify this issue.
In conclusion, we demonstrated that elite basketball athletes have a larger WM volume of the vermian lobules VI and VII (the declive, folium, and tuber) in the cerebellum compared to control subjects, which might reflect a refinement of the connectivity in this region. This result suggests that long-term intensive motor learning evokes WM plasticity in this region of the cerebellum, and that the WM of this region plays a critical role in coordination based on motor learning. The findings of brain plasticity induced by motor learning could contribute to the development of neuro-rehabilitation programs.



 XML Download
XML Download