

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The noninvasive repetitive transcranial magnetic stimulation (rTMS) technique generates an electromagnetic field that can penetrate the skull and brain with minimal attenuation [12]. rTMS produces electrical currents [2] in cortical regions beneath the stimulation coil that can change excitability of these regions and their functionally connected cortical and subcortical structures depending on the intensity and frequency of the stimulation applied. While low frequency rTMS (less than 1-Hz) diminishes the excitability of neuronal cells, high frequency rTMS (at 5-Hz and higher) enhances it resulting in brain activity modulation [34].
The majority of rTMS clinical studies demonstrated improvement of clinical signs beyond therapeutic sessions, which implies functional and structural neuroplasticity in the brain [5678] This phenomenon could be justified by synaptic plasticity, alterations in dendritic spines shape and number or axonal sprouting through long-term potentiation or depression mechanisms [3]. Besides, other structural neuroplasticity mechanisms such as stem cell activation and new nerve cell production might also play a role justifying the outlasting effects of rTMS [9].
Neural stem and progenitor cells (NS/PCs) locate in highly sophisticated niches mainly in neurogenic regions of the brain such as the lateral wall of lateral ventricles (subventricular zone, SVZ) [10] and hippocampus (subgranular zone) [11]. It has been shown that chronic (14-21 days) high frequency rTMS (15-25 Hz) can induce cell proliferation and enhance hippocampal neurogenesis in normal brain and also in animal model of depression [912]. Moreover, high frequency rTMS (7 days, 10 Hz) increased NS/PCs proliferation in the SVZ of animals with focal cerebral ischemia [13].
As mentioned, the majority of studies used high frequency rTMS and to the best of our knowledge there is no study comparing side by side the effects of low and high frequency rTMS on NS/PCs in the literature. Moreover, since there are potential risks administering high frequency rTMS in populations with neurological disorders [1] investigating the effects of low frequency rTMS on NS/PCs properties is needed. This study aimed to evaluate the effects of short (1 week) and long-term (2 weeks) rTMS application at both low and high frequencies (1-Hz and 30-Hz) on NS/PCs proliferation and differentiation in the SVZ of intact adult male mice.
Materials and Methods
Experimental animals
Adult BALB/c male mice (25-30 g, the laboratory animal center of Shiraz University of Medical Sciences, Shiraz, Iran) were housed in groups of five during the course of study under a 12-hour light/dark cycle at room temperature at 25±1℃ with free access to chow diet and water. Animal experiments were conducted in accordance with the Institutional Guidelines and Animal Ordinance (Ethics Committee of Shiraz University of Medical Sciences, Shiraz, Iran). Thirty animals were divided into three groups (n=10) of sham (no stimulation), low frequency (1-Hz) and high frequency (30-Hz) that received rTMS for 1 or 2 weeks.
Repetitive transcranial magnetic stimulation
rTMS was applied using a Neurosoft device (Model 9000 MS, Neurosoft, Ivanovo, Russia) with a 100 mm circular coil. Using the maximum output intensity of the device, the low frequency (1-Hz) group received 150 pulses/day (5-second train, 10-second pause) in 450 seconds and the high frequency (30-Hz) group received 150 pulses/day (1-second train, 5-second pause) in 30 seconds. To reduce stress, animals were gradually acclimated for 10 min/day to a plastic cylinder and habituated to the noise induced by the stimulator for 1 week. During the stimulation period, the animals were restrained in the small plastic cylinder to prevent their movements. The front part of the coil was held at a distance of 2 mm above the animals' head in a horizontal position for the experimental groups and around 10 cm above the animals' skull in a vertical position for the sham groups (450 seconds/day). The animals were treated one session per day for 7 or 14 consecutive days. No signs of seizure or abnormal behavior were noticed in sham or rTMS groups during the length of experiment.
Isolation and expansion of NS/PCs
One day after the last treatment, mice were first anesthetized using 4% isoflurane and sacrificed via cervical dislocation to harvest the brain (animal protocol# 92-6728). A thin layer of tissue containing the SVZ of both lateral ventricles; the rostral part of the brain extending from the olfactory bulb to the crossing of the anterior commissure, was micro-dissected to harvest NS/PCs [1415]. Using a razor blade the harvested tissue was carefully minced into small pieces, and then digested in 0.05% trypsin-EDTA for 5-7 minutes at 37℃. After quenching trypsin-EDTA activity with soybean trypsin inhibitor, the cell pellet was re-suspended in neurosphere medium (500 µl) and mechanically dissociated to achieve a single cell suspension. The cell suspension was spun (110 g, 5 minutes) to remove the supernatant. Finally, the cells harvested from each brain were re-suspended in complete neurosphere medium (5 ml) supplemented with epidermal growth factor (EGF; 20 ng/ml), basic fibroblast growth factor (bFGF; 10 ng/ml), and heparin (2 µg/ml) and incubated in a T25 flask in a humidified incubator with 5% CO2 for 8 days.
Neurosphere forming frequency and size determination
After 8 days of incubation, each culture flask was mounted on the stage of an Olympus inverted microscope (Olympus, Tokyo, Japan) to evaluate the number and size of the resulting neurospheres per brain. Briefly, beginning from the top right corner and ending at the bottom left corner, each flask was thoroughly examined field by field by a researcher blinded to the culture condition and all neurospheres with the size of about 50 µm or bigger in diameter were counted [10] and the numbers were presented as the neurosphere forming frequency per brain. Simultaneously, using a systematic random sampling method, every 30th neurosphere throughout the entire flask was photographed using an Olympus CKX41 digital camera. The diameter of 20-30 representative neurospheres per flask was measured using Stereolite stereological software (Stereolite, SUMS, Shiraz, Iran). The mean neurosphere forming frequency and diameter (as a percentage change relative to the control values) were compared among groups. All data were presented as mean±SEM for each group.
Differentiation of NS/PCs
To analyze the differentiation potential of NS/PCs from each group, the resulting neurospheres were dissociated into single cells and plated in differentiation assay as described before [1617]. Briefly, cells (2-3×105 cells/ml) were first cultured in neurosphere medium containing EGF (20 ng/ml), bFGF (10 ng/ml), and 5% fetal calf serum (FCS) in 24-well plates for 3-4 days in a humidified incubator with 5% CO2. Then, the medium of each well was replaced with a similar medium containing 5% FCS but without growth factors. After 4 days, the cultures were fixed using cold paraformaldehyde (PFA, 4%, 20 minutes at room temperature) and processed for immunofluorescence analysis.
In vitro repetitive magnetic stimulation of NS/PCs
To assess the effect of rTMS on NS/PCs proliferation in vitro, single cells (2.5×105 cells in 5 ml/25 flasks) from dissociated primary neurospheres were plated using complete neurosphere medium supplemented with EGF (20 ng/ml), bFGF (10 ng/ml), and heparin (2 µg/ml) in T25 flasks and incubated in a humidified incubator with 5% CO2 for 8 days (neurosphere formation period). For seven consecutive days, the culture flasks were exposed to sham, 1-Hz, and 30-Hz stimulation. In each session, the 100-mm circular coil of the stimulator was held on the top of each flask applying 1-Hz and 30-Hz stimulation for 450 and 30 seconds, respectively. In sham group, the applicator was held 10 cm above the flasks for 450 seconds in a vertical position. On day 8, the resulting neurospheres from each condition were collected and dissociated into single cells. The number of NS/PCs was quantified for each condition using hemocytometer and expressed as a percentage relative to the sham group.
To assess the effect of rTMS on NS/PCs differentiation in vitro, single cells from dissociated neurospheres were first plated in complete neurosphere medium supplemented with EGF (20 ng/ml), bFGF (10 ng/ml), and 5% FCS in 4-well plates for 3-4 days in a humidified incubator with 5% CO2. Then, the medium of each well was replaced with a similar medium containing 5% FCS but without growth factors and incubated for additional 4 days. During these 4 consecutive days, the culture dishes were exposed to sham, 1-Hz, and 30-Hz stimulation as described earlier. After 4 days, the cultures were fixed using cold PFA (4%, 20 minutes at room temperature) and processed for immunofluorescence analysis.
Immunofluorescence
Representative neurospheres from control and rTMS treated groups were stained as a whole for putative NS/PCs markers [151819] such as nestin and glial fibrillary acidic protein (GFAP). Differentiated NS/PCs cultures were immunostained to determine the percentage of neuronal and glial progeny in cells obtained from different treatment groups. Before proceeding to immunostaining, the samples were fixed using cold PFA (4%, 20 minutes at room temperature) and washed with phosphate buffered saline (PBS) to remove PFA, and then 500 µl of the primary antibody solution containing mouse monoclonal anti nestin (1:500, Millipore, Billerica, MA, USA), mouse monoclonal anti-β-III-tubulin (1:2,000, Promega, Madison, WI, USA) and rabbit polyclonal anti-GFAP (1:500, Dako Cytomation, Carpinteria, CA, USA) in PBST (PBS +0.1% Triton-X) supplemented with 10% NGS, was added to each well of the 24-well plates. After 1 hour of incubation at room temperature, the primary antibody was removed and the samples were washed with PBS. Then, the samples were incubated in a secondary antibody solution containing goat anti-mouse Alexa-fluor 568, goat anti-rabbit Alexa-fluor 488 (1:700, Millipore) and DAPI (1:1,000, Millipore) in PBST supplemented with 10% NGS for 45 minutes at room temperature in the dark. After removing the antibody solution and washing with ample amount of PBS, the samples were analyzed using a fluorescent microscope (Olympus IX-71) equipped with Canon EOS digital camera (Canon, Tokyo, Japan). Representative pictures of each well (10-15 fields/well) were collected. Cells were counted after merging pictures using Adobe Photoshop CS4 and data were presented as a percentage of total cells counted.
Statistical analysis
The results were expressed as mean±SEM and analyzed using GraphPad Prism (version 6.01, GraphPad, San Diego, CA, USA) software. Analysis of Variance (ANOVA) with Newman-Keuls post-hoc test was used to analyze data. The level of significance for all comparisons was set at P<0.05.
Results
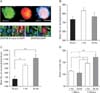
Effects of rTMS on neurosphere forming frequency of the in vivo exposed NS/PCs
The harvested SVZ tissue from all groups proliferated in vitro and formed nestin and GFAP expressing neurospheres after 8 days in culture (Fig. 1A, upper panel). Resulting neurospheres were also capable of differentiating into neurons, astrocytes and oligodendrocytes (Fig. 1A, lower panels) confirming their neural stem cell identity. Neurosphere counting showed that the mean neurosphere number per brain upon one-week rTMS application at 1-Hz and 30-Hz was 832±30.4 and 766±46.3, respectively. These measures were not significantly different from the mean neurosphere number per brain in the 1-week sham group that was 736±26.8 (Fig. 1B). In 2-week treatment paradigm the mean neurosphere number per brain increased significantly from 662.4±68 in the sham group to 968.5±34.3 (P<0.01) and 1,134±73.9 (P<0.001) in the rTMS groups treated with 1-Hz and 30-Hz rTMS, respectively. No significant difference was observed between the two rTMS treatment groups (Fig. 1C). Moreover, statistical analysis revealed that treatment of animals with either 1-Hz or 30-Hz rTMS for 2 weeks resulted in a significant increase in the proliferation of neural stem and progenitors in comparison to the 1-week treatment paradigm (Fig. 1D).
Effects of rTMS on mean neurosphere diameter (size) of the in vivo exposed NS/PCs
Mean neurosphere diameter represents the inherent proliferation capacity of the isolated NS/PCs. Analysis of neurosphere size in 1-week treatment paradigm revealed that the mean neurosphere diameter increased by 10.6±2.8% in 1-Hz group (110±4.23%) and 7.8±0.34% in 30-Hz group (107.8±1.74%), from which only 1-Hz group showed statistically significant difference compared to the sham group (100±1.40%, P<0.05) (Fig. 2A, B). In 2-week treatment paradigm, however, the mean neurosphere diameter (Fig. 2A, C) significantly increased by 14.8±1.94% in 1-Hz group (114.8±4.22%, P<0.05) and 21±2.26% in 30-Hz group (121±4.54%, P<0.01). Furthermore, comparing the mean neurosphere diameter in rTMS treatment groups at 1-week versus 2-week paradigm showed that only 2-week 30-Hz rTMS treatment resulted in significantly larger neurospheres compared to the 1-week 30-Hz rTMS treatment (P<0.05) (Fig. 2D). No significant differences were observed between other treatment groups.
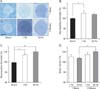
Effects of rTMS on differentiation capacity of the in vivo exposed NS/PCs
Immunophenotyping analysis of representative immunostained pictures (Fig. 3A) of differentiated NSC cultures from different treatment groups revealed that in 1-week rTMS treatment paradigm relative to the sham group, the mean percentages of β-III-tubulin immunoreactive (IR) and GFAP-IR cells were 114±17.53% and 116.8±11.67% in 1-Hz group, and 263±41.94% and 79.77±11.61% in 30-Hz group, respectively (Fig. 3B, E). Statistical analysis showed that only the mean percentages of β-III-tubulin-IR in 30-Hz group were significantly different to the sham and 1-Hz groups (163±27.88% and 149.4±24.41% increase, respectively, P<0.01).
In 2-week rTMS treatment paradigm the mean percentages of β-III-tubulin-IR and GFAP-IR cells relative to the sham group, were 375.5±40.13% and 86.89±1.98% in 1-Hz group, and 307.1±43.23% and 93.44±1.98% in 30-Hz group, respectively (Fig. 3C, F). Statistical analysis revealed that the mean percentage of β-III-tubulin-IR cells in 1-Hz and 30-Hz groups were significantly different to the sham group (273.5±25.43% increase in 1-Hz group, P<0.001 and 207.1±28.53% increase in 30 Hz, P<0.01).
Comparing the mean percentage of β-III-tubulin-IR cells in rTMS treatment groups showed that 1-Hz rTMS treatment significantly increased neuronal cell differentiation at 2-week versus 1-week paradigm (261±22.6% increase, P<0.01) (Fig. 3D). Moreover, the 30-Hz treatment group in 2-week paradigm significantly increased neuronal cell differentiation in comparison to both 1-Hz and 30-Hz rTMS treatment groups in 1-week paradigm (192±25.7% and 43.3±1.29% increase comparing to 1 Hz and 30 Hz, respectively, P<0.05) (Fig. 3D).
The percentage of GFAP-IR cells was significantly higher in 1-Hz rTMS treatment in 1-week paradigm compared to 30-Hz group in 1-week paradigm and 1-Hz group in 2-week paradigm (P<0.05) (Fig. 3G), but no significant differences were noticed between the remaining groups.
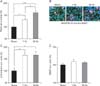
Effects of rTMS on proliferation and differentiation of in vitro exposed NS/PCs
We exposed cultured NS/PCs to sham, 1-Hz and 30-Hz rTMS to investigate whether in vitro application of rTMS could increase their proliferation and change their differentiation capabilities. Counting the total number of cells harvested from different rTMS and sham treated neurosphere cultures after 7 days of repetitive magnetic stimulation demonstrated that compared to the sham stimulation group, NS/PCs number was significantly increased by 32% in 1-Hz (131.4±2.92% P<0.05) and 60% in 30-Hz (160.1±11.14%, P<0.001) groups, respectively (Fig. 4A). Furthermore, the amount of NS/PC proliferation in 30-Hz group was almost 50% more than the 1-Hz group (P<0.05) (Fig. 4A).
Stimulation of NS/PCs in differentiation culture led to a considerable amount of neuronal cell differentiation in both 1-Hz (165±12.56%) and 30-Hz (166.2±16.24%) conditions compared to the sham stimulation condition (P<0.05) (Fig. 4B, C). No statistically significant difference was found between different treatment groups in terms of astrocyte differentiation (Fig. 4D).
Discussion
Despite the use of rTMS as a promising therapeutic tool for many neurological conditions, the exact mechanisms behind rTMS-induced plasticity are still unknown. In present study, for the first time, we evaluated the effect of short and long-term application of both low and high frequency rTMS on the behavior of NS/PCs as one of the key players in neuroplasticity. We found that 2 weeks application of both low (1-Hz) and high frequency (30-Hz) rTMS on adult murine intact brain would increase the proliferation of NS/PCs. This was evidenced by an increase in neurosphere forming frequency and neurosphere size in the rTMS treated animals. In addition, differentiation of the resulting neurospheres showed that rTMS treated NS/PCs in both one-week and two-week paradigms were more neurogenic compared to the sham treated NS/PCs. Moreover, one week in vitro rTMS treatment of NS/PCs with both 1-Hz and 30-Hz increased NS/PCs proliferation and neuronal differentiation.
There are few studies that assessed the effects of rTMS on NS/PCs proliferation and differentiation in vivo [91213] employing bromodexoiuridene (BrdU) labeling method. Ueyama et al. [9] showed that after 14 days of applying high frequency (25-Hz) rTMS (1,000 pulses/day), cell proliferation in dentate gyrus of hippocampus increased of which most of cells expressed neuronal phenotype. In a similar study, Feng et al. [12] demonstrated that application of high frequency (15-Hz) rTMS (1,000 pulses/day) for 21 days in a chronic rodent model of depression increased both hippocampus cell proliferation (increased neurogenesis) and brain-derived growth factor (BDNF) protein level in the dentate gyrus of the hippocampus. In a rat model of focal cerebral ischemia, Guo et al. [13] demonstrated that the application of high frequency (10-Hz) rTMS (300 pulse/day) for 7 days significantly increased proliferation of NS/PCs in the SVZ of the lateral wall of lateral ventricle. We applied both low (1-Hz) and high (30-Hz) frequency rTMS (150 pulse/day) for 7 and 14 days and used the neurosphere assay and an in vitro differentiation method to evaluate NS/PCs behavior. Overall, our results are in line with the findings of previous studies by Ueyama et al. [9] and Feng et al. [12]. Unlike the study by Guo et al. [13], we could not see any significant proliferation of NP/SCs upon one week rTMS application in terms of neurosphere forming frequency unless a slight increase in neurosphere size in 1-Hz rTMS treatment group. This discrepancy could be attributed to cerebral ischemia model that by itself can cause NP/SCs proliferation [20]. It appeared that application of high frequency rTMS for 7 days would potentiate NP/SCs proliferation and neurogenesis in ischemic but not intact brain, as they did not include any results suggesting the same effects in the intact brain.
Interestingly, our findings showed that low frequency rTMS is as equally effective as high frequency rTMS in recruiting NS/PCs into proliferation and increasing their neuronal differentiation. We found a trend of increase in neurosphere number and diameter in both low and high frequency rTMS at 1-week treatment paradigm but only the change in neurosphere diameter at low frequency rTMS reached statistical significance. This implies that rTMS even at low frequency upon one-week application could result in at least a subtle increase in NS/PCs proliferation. To have a more pronounced effect, a longer (2 weeks) application time is needed for both the low and high frequency rTMS. Given that the treatment with low frequency rTMS is as effective as high frequency rTMS, this provides us with a safer therapeutic option that is tolerated better and imposes fewer risks comparing to the high frequency rTMS.
Possible mechanisms behind the increased NP/SCs proliferation and neuronal differentiation upon rTMS application have not yet been very well characterized. Up-regulation of neural stem cell growth promoting factors such as BDNF was shown to be key players for hippocampal cell proliferation and neuronal differentiation upon rTMS treatment [12]. BDNF also increases NS/PCs proliferation and new nerve cell production in the SVZ [21]. Alternatively, the release of neurotransmitters by axonal terminals innervating the SVZ could be another contributing factor to NS/PCs proliferation in the SVZ [22]. In fact, several studies have shown that the SVZ is rich in nerve endings originating either from the local neural circuitry such as GABAergic neurons of the adjacent striatum [232425] or from distant brain regions such as dopaminergic neurons of the substantia nigra, ventral tegmental area [2627], and serotonergic neurons of the raphe nuclei [28]. GABA is an inhibitory neurotransmitter that maintains a balance in proliferation and modulates the quiescence of NP/SCs in SVZ [24]. Dopamine [2930] and serotonin [28] have proven positive effects on NS/PCs proliferation in the SVZ. Hence, we can speculate that activation of different neurotransmitter system by rTMS may actually affect the SVZ stem cell niches with final net result of increased cell proliferation in rTMS treated animals. Apart from these, our in vitro study showed that the exposure of the cultured NS/PCs to both low and high frequency repetitive magnetic stimulation would increase cell proliferation and neuronal differentiation (Fig 4). This implies that electromagnetic fields per se would affect cell proliferation and differentiation. In support of this findings, Sun et al. [31] showed that application of pulsed electromagnetic fields on human bone marrow mesenchymal stem cells for 8 hours per day during culture period would result in around 50% more viable cells at 24 hour after plating. In addition, Piacentini et al. [32] demonstrated that extremely low-frequency electromagnetic fields exposure promote neuronal differentiation of NS/PCs by upregulating Ca(v)1-channel expression and function [32]. Overall, all these mechanisms have the potential to mediate rTMS effects on NP/SCs; however, further investigations are needed to find the exact interactions behind rTMS effects on NP/SCs proliferation and neurogenesis.
Conclusion
This study confirms that both low and high frequency rTMS increase NS/PCs proliferation and neurogenesis in the SVZ of the adult intact murine brain. This findings encourages the application of using low frequency rTMS as a safe and equally beneficial neuromodulator as the high rTMS when the stimulation of neural stem cells are intended to promote regeneration in diseased brain conditions. Furthermore, due to the positive effect of rTMS on both NS/PCs proliferation and neuronal differentiation, rTMS could be suggested as a tool to establish more neurogenic neural stem cell lines for therapeutic purposes, although more detailed cellular and molecular studies are needed to reach ideal settings.

 XML Download
XML Download