

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Ischemia-reperfusion injury (IRI) is an inevitable consequence of renal transplantation and appears in early kidney transplant dysfunction [1, 2]. IRI results in acute kidney failure, which is a major cause of delayed graft function and leads to allograft rejection [3].
The pathogenesis of IRI is not completely understood but has been demonstrated to be related to inflammatory responses and consequent renal cell death. Infiltrating leukocytes damage the renal membrane and produce various pro-inflammatory cytokines. Thus, excessive immune responses result in renal cell death and cause allograft failure. In order to suppress inflammatory responses and increase the rate of transplant success, calcineurin inhibitors (CNIs) are widely used as immunosuppressant [2, 4].
Calcineurin is a serine/threonine phosphatase whose activity is required to mediate various intracellular calcium signaling pathways [5]. In response to various stimuli, calcineurin dephosphorylates transcription factors such as the nuclear factor of activated T-cells (NFAT) [6]. Dephosphorylated NFAT induces the release of interleukin-2, -3 and -4, interferon-γ, and tumor necrosis factor-α (TNF-α) and results in various immune responses [6]. Calcineurin, in particular, plays an essential role in T-cell activation [6] as a key component regulating T lymphocyte signal transduction [7, 8, 9, 10]. In myocardial ischemia, calcineurin has been shown to regulate both pro- and anti-apoptotic factors [7, 11]. Because of the properties of calcineurin, CNIs are widely used to suppress immune responses. FK506 is a well-known CNI [5, 11] that inhibits calcineurin activity though binding to the FK binding protein (FKBP) domain [11, 12, 13, 14]. However, FK506 has shown nephrotoxicity when used in excess, inducing renal cell death [10, 12].
In this study we examined the effects of FK506 in a renal IRI mouse model. First, we showed calcineurin expression induced by IRI and determined a minimum concentration of FK506 that is effective in reducing calcineurin activity without kidney damage. Calcineurin is composed of calcineurin A and calcineurin B subunits. FK506 has been shown to bind to FKBP12, which is a protein distinct from calcineurin A. The FK506-FKBP12 complex inhibits calcineurin activity [12, 15, 16]. Further, we suggest that the role of FK506 in the nucleus is different from its role in the cytoplasm. Nuclear calcineurin has been reported in myocardial hypertrophy [8, 17, 18, 19, 20]. According to these studies, calcineurin senses intracellular Ca2+ by undergoing a conformational change in its C-terminal autoinhibitory (AI) domain upon binding to Ca2+. Targeted proteolysis of the calcineurin AI domain by calpain increases calcineurin activity, which contributes to the translocation of NFAT to the nucleus [17]. In cardiomyocytes, calpain stimulation by angiotensin II causes the proteolytic cleavage of calcineurin [21]. The cleaved calcineurin functions in the nucleus with increased phosphatase activity. Once activated, calcineurin dephosphorylates NFAT, translocates to the nucleus, and activates NFAT-mediated transcription-inducing genes related to pro-inflammatory cytokines. Thus, we assume that FK506, a typical inhibitor of calcineurin, attenuates the cleavage of calcineurin and consequent inflammatory responses. In addition, we clearly demonstrated the expression of calcineurin and related inflammatory molecules through immunohistochemistry, which has been difficult to show directly in the kidney in IRI models in previous studies. Here, we re-evaluated FK506 as a new therapeutic agent as an anti-inflammatory factor for treating IRI as well as other, related diseases.
Materials and Methods
Animals
Male C57BL/6 mice were purchased from Central Laboratory Animal, Inc. (Seoul, Korea). Mice used in all experiments were 12 weeks old. These mice were housed in a specific pathogen-free facility with appropriate temperature and humidity and allowed free access to food and water. The mice for this study (GNU-121108-M0046) were approved by the Institutional Animal Care and Use Committee at Gyeongsang National University.
IRI mouse model
The mice were anesthetized by intramuscular injection of Zoletil (30 mg/kg, Virbac Laboratories, Carros, France) and Rompun (10 mg/kg, Bayer Korea, Seoul, Korea). The temperature of the anesthetized mice was adjusted to near 37℃ using a warming light. Animals were allowed to self-breathe room air. Both kidneys were exposed by a longitudinal incision, and the renal pedicle was occluded by atraumatic microaneurysm clamps for 25 minutes to induce ischemia. After 25 minutes, the clamp was removed for reperfusion. The sham group was performed by the same surgical method except that the clamping was not applied. FK506 was injected intraperitoneally 30 minutes before surgery. The concentrations were 0.01, 0.1, 1, 10, and 100 µg/kg. Twenty-four hours after surgery, the mice were anesthetized and perfused transcardially with heparinized saline. One kidney was removed and frozen in liquid nitrogen immediately and the other kidney was fixed in 4% neutral buffered formalin. Blood samples were collected to measure creatinine levels from the heart before perfusion.
Measurement of creatinine levels
Blood samples from the heart were collected in glass evacuated blood collection tubes (Greiner Bio-one, Frickenhausen, Germany) and centrifuged at 4℃. Blood serum analysis was requested for measurement of creatinine level at the Korea Green Cross Corp. (Yongin, Korea).
Immunohistochemistry
After fixation, kidney tissues were paraffin-embedded and cut into 5-µm-thick longitudinal sections. After deparaffinization, the slide samples were incubated with appropriate primary antibodies as follows. Antibodies against calcineurin (BD Bioscience, San Jose, CA, USA), neutrophil infiltration protein Ly6B.2 (clone 7/4, AbD Serotec, San Diego, CA, USA), TNF-α (Abcam, Cambridge, UK), caspase-3 (Asp175, Cell Signaling Technology, Danvers, MA, USA), NFATc1 (NFAT2, Santa Cruz Biotechnology, Santa Cruz, CA, USA), and intercellular adhesion molecule-1 (ICAM-1; Sino Biological Inc., Beijing, China) were used. Sections were incubated with primary antibody at 4℃ overnight. After washing three times with 0.1 M phosphate buffered saline (PBS), sections were incubated with the biotin tagged an appropriate secondary antibody at room temperature (RT) for 1 hour. After washing three times with 0.1 M PBS, sections were incubated in avidin-biotin-peroxidase complex solution (Vector Laboratories, Inc., Burlingame, CA, USA) and developed with diaminobenzidine substrate kit (Vector Laboratories, Inc.). After dehydration, sections were mounted and visualized with a BX50 microscope (Olympus, Tokyo, Japan), and digital images were captured. Captures were randomly distributed on 5 areas of all field and represented as high power field using Image J.
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
After fixation, kidney tissues were paraffin-embedded and cut into 5-µm-thick longitudinal sections. After deparaffinization, sections were treated with 3% H2O2 in methanol for peroxidase quenching, permeabilized with 0.1% Triton X-100, and labeled using the In Situ Cell Death Detection Kit (Roche Applied Science, Mannheim, Germany). The fluorescence images were captured with a BX50 microscope (Olympus). Statistical analysis was performed using the Image J.
Tissue fractionation and western blot
Frozen kidney tissues were fractionated using the Nuclear/Cytosol Fractionation Kit (BioVision Inc., Milpitas, CA, USA) according to the manufacturer's protocol. Protein concentration was determined using a bicinchoninic acid kit (Pierce, Rockford, IL, USA). Isolated nuclear and cytoplasmic extracts were separated by 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. The membranes were incubated in blocking solution (3% bovine serum albumin in TBS containing 0.1% Tween-20) at RT for 1 hour and then with the following primary antibodies. Antibodies against calcineurin (BD Biosciences), calpain 1 (H-65), the cytosol marker PRX II (Santa Cruz Biotechnology), and the nuclear protein p84 (Abcam) were used. The protein bands were visualized by an enhanced chemiluminescence western blot detection system (Amersham Pharmacia Biotech, Piscataway, NJ, USA). Digital images were capture with an ImageQuant LAS-4000 Imager (GE Healthcare Life Sciences, Buckinghamshire, UK) and analyzed by Image J.
Statistical analysis
Statistical analysis was performed using SPSS version 21 (IBM Co., Armonk, NY, USA). Results are represented as mean±SD. Analysis was based on three independent experiments, each of which included 4 mice per group. P-values were determined through one-way ANOVA, and less than 0.01 were considered statistically significant.
Results
Renal IRI mouse model and changes in creatinine levels
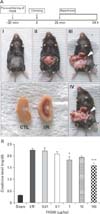
FK506 is a widely used immunosuppressant in most organ transplantation procedures [1, 22]. We examined the effects of FK506, which is known to inhibit calcineurin activity, and consequent renal cell death in renal ischemic injury. To address this, we established an in vivo renal IRI mouse model. FK506 pre-conditioning was performed by injection 30 minutes before surgery. By clamping renal arteries, we confirmed the color of kidney became darkened, and reperfused by unclamping. After 24 hours, the kidneys were surgically removed, and we found that the injured kidney displayed an infarct in the outer medulla compared to the control kidney, as shown in Fig. 1A.
We first examined the effect of FK506 on renal function in our IRI model. To determine the dose-dependent effect of FK506, the mice were treated with FK506 at various concentrations, as indicated. To assess renal function, we measured serum creatinine levels, elevations of which are indicative of renal filtration dysfunction. We confirmed that creatinine levels were significantly increased more than 5-fold in ischemia/reperfusion (I/R) mice compared to the sham control mice. FK506 caused reductions in creatinine levels at concentrations of 1 µg/kg or greater (Fig. 1B), showing that FK506 can improve renal function. In addition, FK506 dose-dependently reduced infarction of outer medulla (data not shown).
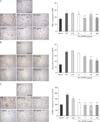
FK506 reduced the expression of calcineurin and NFAT2 in renal IRI mice
Because FK506 is a well-known inhibitor of calcineurin [5], we examined changes of calcineurin expression after FK506 treatment by performing immunohistochemistry (Fig. 2A). As previously reported [13, 23], calcineurin expression was observed in the proximal and distal tubules and was found to be highly expressed in the outer medulla after IRI. We confirmed that calcineurin expression was significantly increased by I/R and decreased by FK506 treatment at concentrations of 0.1 µg/kg or higher. The sham control kidneys had low levels of calcineurin. Therefore, these results demonstrate that FK506 is effective in reducing calcineurin expression.
NFAT is an important transcription factor that directly regulates the expression of pro-inflammatory cytokines in T and B lymphocytes during adaptive immune responses [24, 25, 26, 27, 28]. Calcineurin dephosphorylates NFAT2 in the cytoplasm, and the dephosphorylated NFAT2 translocates to the nucleus. Nuclear NFAT regulates transcriptional activity of target genes. Thus, by immunohistochemistry we measured changes in NFAT2 expression following FK506 treatment (Fig. 2B). As expected, NFAT2 was dramatically induced in I/R mice about 3-fold compared to the sham control mice and reduced by FK506. Here, we showed that reduced expression by FK506 could negatively regulate the calcineurin-NFAT2 signaling. Eventually, FK506 mitigate the inflammation of kidney outer medulla by excessive expression of calcineurin-NFAT2.
FK506 reduced apoptotic cell death, pro-inflammatory cytokine expression, and neutrophil infiltration protein in renal IRI mice
To investigate whether FK506 has a protective role in renal cell death, we performed the TUNEL assay. Apoptotic nuclear staining was evident in renal IRI mice and dramatically reduced by FK506 treatment to a level similar to that in sham control mice (Fig. 3A). Then, we examined levels of cleaved caspase-3 as an apoptotic marker by immunohistochemistry (Fig. 3B). In particular, a FK506 dose of 0.1 µg/kg, which is 1,000-fold lower than the widely used dose, was sufficient to clearly reduce caspase-3 expression.
Then, we examined the inflammatory signaling pathways that could be affected by FK506 and measured the expression of TNF-α, a pro-inflammatory cytokine, by immunohistochemistry (Fig. 4A). The number of TNF-α-positive cells was significantly increased, and the immunoreactivity was intense in renal IRI mice compared to the sham control mice. TNF-α expression was strong in the proximal and distal tubules around the glomerulus, and FK506 particularly reduced its expression in distal tubules of the cortex and tubules in the outer medulla. We also found that calcineurin and TNF-α were expressed in identical regions.
Stimuli such as neutrophil infiltration evoke excessive expression of pro-inflammatory cytokines and are also known to mediate IRI [29]. Thus, we investigated whether FK506 affects neutrophil infiltration by immunohistochemistry (Fig. 4B). Staining of neutrophil infiltration protein Ly6B.2 was apparent in the outer medulla of IRI mice and increased about 3-fold compared to the sham control mice. In addition, neutrophil infiltration was apparent in the interstitium, where blood vessels are mostly distributed, of the I/R mice. However, the level of neutrophil infiltration in the interstitial tubules was greatly reduced after FK506 treatment. These results suggest that FK506 exerts its anti-apoptotic function in IRI through reducing inflammatory cytokines and neutrophil infiltration.
FK506 reduced extracellular matrix accumulation in renal IRI mice
Excessive pro-inflammatory cytokines can induce accumulation of the extracellular matrix (ECM), which is known to cause renal diseases such as fibrosis [30] and diabetic nephropathy [7, 23, 30]. We observed that FK506 treatment decreased TNF-α, expression and therefore would suppress ECM accumulation. Thus, we examined the expression of ICAM-1. ICAM-1 is a well-known ligand for integrin LFA-1 and regulates the recruitment of inflammatory leukocytes. As shown in Fig. 4C, we found that ICAM-1 expression was dramatically increased in I/R mice and FK506 treatment suppressed ICAM-1 expression to a level similar to that in the sham control mice. ICAM-1 was normally expressed in the interstitium of the kidney, where blood vessels are distributed, as shown in the sham mice. ICAM-1 expression was further induced in the glomerulus, where capillaries are distributed, in the I/R mice. Thus, we found strong ICAM-1 staining in the thickened interstitium. FK506 at a dose of 1 µg/kg was sufficient to reduce the expression of ICAM-1.
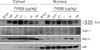
FK506 reduced calpain-mediated calcineurin activity
We next investigated the activity of calcineurin in the nucleus after renal IRI. In cardiomyocytes, calcineurin is regulated by calpain-mediated proteolysis [8, 17]. We assumed that calcineurin activity might be regulated in a similar way in the kidney and play a crucial role during renal IRI. Thus, we prepared cytosolic and nuclear extracts from kidney of IRI or sham control mice and examined calcineurin expression patterns (Fig. 5). We detected the 48-kDa cleaved form of calcineurin along with the 58-kDa full-length form of calcineurin. The cleaved form of calcineurin was found only in IRI mice and was presumably processed by calpain. This result showed that FK506 decreased calcineurin expression and further reduced levels of the cleaved form of calcineurin. FK506 did not appear to affect calpain activity directly; no substantial changes were observed in nuclear calpain levels. This result was confirmed by immunohistochemistry using calpain-1 antibody (data not shown). We suggest that FK506 reduces the activity of calcineurin in both the nucleus and the cytoplasm and that particularly in the nucleus FK506 has a cytoprotective effect by inhibiting NFAT2-mediated transcriptional activity.
Discussion
Renal IRI involves a complex pathophysiologic mechanism, and calcineurin has been proposed as a major molecular mediator of IRI [13]. It is a Ca2+/CaM-dependent protein phosphatase and is an important Ca2+ effector promoting necrosis and apoptosis [23, 31, 32, 33, 34, 35]. Calcineurin activity is predominant in the cytoplasm. However, we suggest here that calcineurin has activity in the nucleus, particularly in a renal I/R model. Previously, its role in the nucleus has been discussed during ischemia in other tissues. In this study, we reinvestigate FK506, a well-known CNI, and confirmed its role in reducing expression of the pro-inflammatory cytokine TNF-α and decreasing neutrophil infiltration. Despite its beneficial roles in reducing inflammatory responses, FK506 exhibits adverse effects when administered in high doses [10, 22, 36]. Therefore, we sought the lowest effective dose of FK506 and demonstrated that 1 µg/kg is sufficient, which is 1,000 times less than the normal dose (1 mg/kg).
We thought that FK506 could improve renal function. Thus, we measured serum creatinine level. However, serum creatinine level is not sufficiently explained renal function. To assess renal function, glomerulus filtration rate is required. Therefore, we could be restricted to only anti-inflammatory effects of FK506. Fig. 1B showed serum creatinine level decreasing by FK506 treatment. The creatinine level was clearly reduced, but not significant than pro-inflammatory cytokine level. The creatinine level is closely linked to the renal failure and impairment. However, creatinine level is influenced by various factors such as status of muscle, protein uptake and high dietary of meat [37, 38]. Thus, serum creatinine level does not need to be exactly same with the reduction of the inflammatory cytokine.
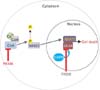
Then, we investigated the role of calcineurin in the nucleus. To our knowledge, this is the first study of nuclear calcineurin in renal IRI. We separated the cortex and medulla of the kidney and performed western blot analysis. We clearly showed a cleaved form of calcineurin in addition to full-length calcineurin. Calpain is known to catalyze the cleavage of calcineurin in the nucleus, which promotes NFAT2 transcriptional activity [17]. We speculated that the inhibitory effect of FK506 is not confined to the cytoplasm as previously shown, but may have a critical role in the nucleus as our results suggest. FK506 may block calpain-mediated calcineurin activity in the nucleus (Fig. 6). FK506 did not directly affect calpain activity, but diminished immune responses presumably by inhibiting the action of cleaved calcineurin. We suggest here that FK506 decreases calcineurin activity not only in the cytoplasm, but also in the nucleus. Furthermore, this dual effect may suppress overall inflammatory responses induced by renal IRI.
Although the FK506 effect on the medulla was not investigated in this study, we detected the cleaved form of calcineurin in the cytoplasm of cells of the cortex (Fig. 5) and medulla (data not shown). However, the cleaved form of calcineurin in the nucleus has previously been shown not to translocate back to the cytoplasm of cardiomyocytes [8]. Thus, the role of the cleaved form of calcineurin in the cytoplasm of cells of the renal cortex and medulla is particularly interesting. We performed immunohistochemistry to detect calcineurin, which revealed that calcineurin was expressed in the tubules of the medulla. We speculated that this is related to the presence of the cleaved form of calcineurin in the cytoplasm. Therefore, the cleaved form of calcineurin is worth studying further. FK506 is a well-known immunosuppressant that is commonly used during kidney transplantation to reduce renal IRI because it is known to have anti-inflammatory activity. It has therapeutic potential in other diseases associated with inflammatory responses. We propose that it can be applied to auto-immune diseases such as atopic dermatitis and rheumatoid arthritis and even as an anti-cancer drug [26, 39] because these diseases are closely related to excessive inflammation, including leukocyte infiltration and ECM accumulation [26, 40, 41, 42]. Recently, several studies on the anti-cancer effects of FK506 have been reported [39]. In conclusion, we revisited the effect of FK506 as an anti-inflammatory factor in renal IRI and suggest it inhibits calcineurin activity in both the cytoplasm and the nucleus by inhibiting NFAT2-mediated transcriptional activity.


 XML Download
XML Download