

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The future terminal portion of the inferior vena cava (IVC), is formed by joining of the venous pole of the heart and the omphalo (vitelline)-umbilical venous trunks [1, 2]. At 4 weeks of gestation, the subdiaphragmatic anastomosis connecting the cranial portions of the two vitelline veins above the hepatic primordium, often called the subdiaphragmatic vestibulum, receives blood from the liver [3, 4, 5], suggesting that the two vitelline vessels may correspond to the right and left hepatic veins. At 5 weeks of gestation, the right part of the subdiaphragmatic anastomosis enlarges and fuses with the cranial parts of the hepatic veins to form the common hepatic vein. However, in those previous descriptions, we hardly made an image of the topographical anatomy of the early IVC terminal in and around the heart. One of the reasons was found in absence of the sinus venosus morphology in explanation of early development of the IVC.
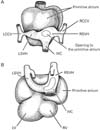
Usually, histological demonstration of the human sinus venosus did not include the IVC (e.g., Blom et al. [6]). Moreover, when the researchers wished to show the IVC terminal, photographic demonstrations of the venous pole of the heart were consistently provided by frontal or sagittal sections [1, 2, 7, 8]. Therefore, it seemed to be still unclear which structure is adjacent to the IVC terminal: the right atrium, left atrium or both. In particularly, the topographical relation of the left and right sinus venosus horns (RSVHs) with the IVC might not be demonstrated in contrast to rather simple diagrams in textbooks of human embryology (Fig. 1): do the three venous elements join together to open the right atrium? In the present study, according to Anderson et al. [1] and Sizarov et al. [2], we called the RSVH to a terminal course of the IVC after joining the left sinus venosus horn. Likewise, in the larger specimens without the left sinus horn, we will use the term for the upper continuation of the IVC in the upper side of the coronary sinus opening. Consequently, in the present study, we attempted to examine serial horizontal sections including not only the heart but the liver to reveal topographical anatomy of the embryonic venous pole of the heart.
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). Horizontal serial paraffin sections stained with hematoxylin and eosin had been obtained from 15 human embryos of crown-rump length (CRL) 9-31 mm, corresponding to gestational ages of approximately 6-7 weeks or Carnegie stage 14-16 [2, 8]. All specimens were part of the large collection kept at the Embryology Institute, Universidad Complutense, Madrid, and were products of miscarriages and ectopic pregnancies managed at the Department of Obstetrics at the university. The study protocol was approved by our university ethics committee (No. B08/374). Our previous studies [9, 10] used heads, necks and abdomens of the same embryos without any anomalies.
Results
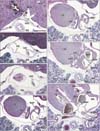
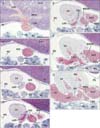
We identified the left sinus venosus horn in 5 of the 15 specimens: the specimen size or CRL was 9 mm, 10 mm, 11 mm, 14 mm, and 15 mm (Figs. 2, 3). Conversely, in specimens larger than 16 mm CRL, we observed the coronary sinus draining into the right atrium (Figs. 4, 5, 6). The IVC terminal and its upper continuation (i.e., the RSVH) consistently provided a thick, continuous tube-like structure. Much or less extent, the IVC was not attached to any part of the heart and liver (Figs. 2C, 3B, 4B, 4C, 5C, 6A). Especially in 3 of the 15 embryos, a part of the course was absolutely free in the pericardial cavity (Fig. 5C). In 9 of the 15 embryos (Figs. 3B, 3C, 4F, 5D, 5E, 6C, 6D), the IVC was connected with the developing right pulmonary vein by a mesentery-like fibrous tissue. The wall of the IVC was thin near the liver, but, during its upward course, muscular elements of the sinus venosum or right atrium appeared to join the IVC (Figs. 3C, 4D). The caudal end of the sinus was separated from the IVC and free in the pericardial space in 2 embryos (Fig. 3B).
The right atrium was consistently attached to the ventral aspect of the IVC and RSVH along a relatively long course (more than 1 mm in Fig. 6). The right auricle or appendage was seen ballooning from the cranial part of the atrial wall (Figs. 2J, 2K, 3E, 3F, 4G, 6E, 6F). Notably, depending on regression of the left sinus venosus horn, instead, the left atrium appeared to the left side and adjacent to the IVC (Figs. 5D, 6C). The left atrial cavity was separated by the left sinus valve from the RSVH (Figs. 5F, 6E). Thus, the left sinus valve appeared to be the interatrial septum, but we discriminated the valve from the septum according to observations of the serial sections. The septum primum was seen in the ventral and left sides of the left sinus valve (Figs. 2K, 3G, 3H), while a candidate of the septum secundum was seen near and above the ventral attachment of the left sinus valve (Fig. 6G). Before development of the septum primum, the left sinus valve was a definite septal structure between the RSVH and left atrium. The site at which the bilateral pulmonary veins joined was consistently separated from the RSVH by the left sinus valve. When the pulmonary veins reached the heart, the left sinus valve became evident.
The IVC and RSVH were mostly or partly surrounded by the left and right venous valves, but the opening of the RSVH to the atrium was located, in 14 of the 15 specimens, on the far caudal side of the opening of the right common cardinal vein, with the distance between the caval vein terminals ranging from 0.3-0.7 mm. Thus, almost always, both the IVC and the future superior vena cave were not seen in the same section. One exception is shown in Fig. 2, in which both caval veins were joined in a panel (Fig. 2H). Because the sinus valve was better developed on the right than on the left side, the RSVH in 8 specimens opened to the left atrium or pulmonary vein before (in the lower side of) opening to the right atrium (Figs. 3G, 6G). In one single specimen, the RSVH opened bilaterally in a single section (Fig. 5G). Because the IVC and RSVH were absolutely continuous, we did not find a reason why we had to discriminate the RSVH from the IVC in the upper side of the coronary sinus opening. Therefore, the IVC (not the RSVH) was likely to open the left atrium in the lower side of the right-sided opening because of the left sinus valve much smaller than the right one.
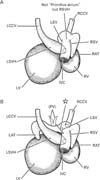
Consequently, rather than the future superior cava, the IVC and RSVH were a major provider of a right-sided atrial lumen at 6-7 weeks. The observations were summarized in Fig. 7.
Discussion
We found that the right atrium extended far caudally along the IVC at stages in which a lumen of the IVC and RSVH occupied in most or major parts of the right atrial structure. In contrast to a fold-like scheme provided by Blom et al. [6], the right sinus valve absolutely separated most of the long IVC-RSVH course from the right atrium. Thus, we suspected almost complete absorption of the right sinus valve to provide the smooth-walled atrium. Instead, another possibility seemed to be likely that, in the later stage, the growth or elongation of the IVC is much smaller than other parts of the right atrium. The IVC was often fixed to the right pulmonary vein by a mesentery-like fibrous tissue. The IVC developing in the pericardial cavity earlier than the pulmonary vein development may provide a traction to make the so-called pulmonary pit for the joining of the later veins. Actually, a developing vessel is likely to make a thick fascial sheath in human fetuses [11]. This fibrous fixation may explain the pathogenesis of the scimitar anomaly [12].
The usual diagram including the large primitive atrium connected with the sinus venosus (Fig. 1) (e.g., Sadler [13], Moore and Persaud [14]) seemed to be unlikely because the atrium extended inferiorly along the IVC. The IVC and RSVH were as thick as the early atrial cavities (Fig. 7). Moreover, the left atrium was likely to be adjacent to the IVC in the present specimens of CRL 9-31 mm. The simplest explanation was found in a stage-dependent change, with the left atrium developing caudally to separate the sinus venosus or coronary sinus from the IVC and RSVH during different stages. However, the typical figure of the left and right sinus valves surrounding the IVC in the right atrium, such as shown in previous studies [1, 7], seemed to be a rather rare morphology or a product chosen in frontal or sagittal sections. Although the "stages" are believed to determine any significant difference in topographical relation in the heart, depending on individual variations, the left sinus valve was likely to separate the IVC from the left atrium. Likewise, we found that, along the long course, the IVC was attached to the dorsal aspect of the right atrium. In adult hearts, the IVC wall attaches to the right atrium at a wide area, while the former attaches to the left atrium at the right posterior margin. We considered that these attachment sites were derived from the right and left sinus valves, respectively. This hypothesis did not contradict with the incorporation theory of the sinus venosus into the right atrial wall [7].
In conclusion, instead of the so-called primitive atrium (Fig. 1), the IVC and RSVH were a major provider of a right-sided atrial lumen at 6-7 weeks (Fig. 7). Variations in topographical anatomy around the IVC, especially of the sinus valves, might not always depend on the stages but partly in individual differences.



 XML Download
XML Download