

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Although, the avian and mammalian testicular architecture is nearly similar, the epididymal tissue is quite different. The avian epididymal region comprises rete testis, proximal and distal efferent ductules (DED), a short connecting duct, and ductus epididymis. However, the mammalian epididymis is traditionally divided into initial segment, head, body, and tail [1, 2, 3, 4, 5, 6]. In addition, most of the avian species does not have accessory sex glands which add secretory products to the semen, but does have secretory cells in the epithelium of the excurrent ducts [7]. Interestingly, the epididymis plays an important role in the re-absorption of testicular fluid [8]. Although sperm traverse this region of the male reproductive tract rapidly, these ducts resorb nearly 90% of the testicular plasma output before the sperm are stored for an extended period in the ductus deferens [8]. The epididymal region of the male reproductive tract is therefore essential for sperm maturation, and dysfunction of this region results in infertility [9, 10, 11].
S-100 proteins, named for their solubility in a 100% saturated solution of ammonium sulphate at neutral pH [12]. It is belong to a group of closely related, small, acidic, water-soluble, Ca2+-binding proteins [13, 14]. A great body of evidence suggests that S-100 could be viewed as a multifunctional subfamily of Ca2+-binding proteins of the EF-hand type. A large number of diverse functions is attributed to S-100 proteins, ranging from calcium-buffering through intracellular (e.g., modulation of enzyme activities, energy metabolism, motility, and secretion) and nuclear (e.g., transcription and apoptosis) functions to extracellular activities (e.g., secretion, neurite extension, and chemotaxis) [15, 16, 17, 18, 19]. Despite all of these proposed functions, the exact biological role of this protein in the testis and epididymis is not yet known. The localization of S-100 was investigated in the testis and epididymis of several mammalian species including bovine [20, 21, 22, 23], sheep [21], rat [21, 24, 25], cat [21, 26], ram, boar, horse, dog [20, 21], buffalo [27, 28], monkey [29], and human [24, 30]. Conversely, one report concerning the S-100 localization in the testis and epididymis of adult White Peking ducks is, to our knowledge, available [31]. Therefore, the present study was conducted to demonstrate S-100 in the testis of adult male of fowls (Gallus gallus domesticus), Sudani ducks (Cairina moschata), pigeons (Columba livia), and rabbits (Oryctolagus cuniculus).
Materials and Methods
The adult, sexually active male birds and rabbits used in this study were purchased locally and maintained under recommended husbandry conditions. Our experiments were carried out according to the institutional ethical committee of the Mansoura University, Egypt.
Tissue preparation
The testes and epididymis of roosters (n=5), male Sudani duck, a local breed of Muscovy found in Egypt (n=3), pigeons (n=5) and rabbits (n=3) were obtained after slaughtering and evisceration of these birds and animals. Small samples of the testicular tissue and their associated epididymis (0.5-1 cm3) were fixed in Bouin's solution for 24 hours. The Bouin's fixed samples were extensively washed in 70% ethanol to remove the fixative before the subsequent steps of tissue processing. Thereafter, the tissue samples were dehydrated in ascending grades of ethanol (70%, 80%, 95% and absolute), cleared in xylene and embedded in paraffin wax using standard techniques. Sections (5 µm) were cut on Leitz microtome and mounted on both coated and uncoated slides.
Immunohistochemical staining
For the detection of S-100, a rabbit polyclonal primary antibody against cow S-100 proteins (Code-Nr. Z 0311, Dako, Hamburg, Germany) was used. Antigen localization was achieved using the avidin-biotin complex technique [32]. Briefly, 5-µm sections of paraffin-embedded tissue were dewaxed, rehydrated, and rinsed in phosphate buffered saline (PBS) pH 7.4 (3×5 minutes). Endogenous peroxidase was blocked by soaking the sections in 3% v/v hydrogen peroxide/distilled water for 10 minutes at room temperature followed by washing them under running tap water for additional 10 minutes. Subsequently the slides were rinsed in PBS pH 7.4 (2×5 minutes). Non-specific antibody binding was minimized by covering the slides with a serum-free protein blocking reagent (Dako, Hamburg, Germany) for 10 minutes at room temperature. Sections were then incubated for 30 minutes at room temperature with primary antibody diluted 1:400 in antibody diluent (Dako, Hamburg, Germany).The slides were subsequently soaked in PBS pH 7.4 (2×5 minutes) followed by incubation with diluted (1:300 in PBS) biotinylated secondary antibody (pig anti-rabbit IgG). Bound antibodies were visualized using a streptavidin-biotin peroxidase complex kit and diaminobenzidine (Dako, Munich, Germany). All incubations were performed in a humidified chamber. Sections were left unstained or counterstained in Mayer's haematoxylin, dehydrated, and mounted with DPX (Sigma, Munich, Germany). Negative controls were performed by omission of the primary antibody. For scoring the S-100 immunoreactivity, image J program v.1.44 was applied.
Results
In this study a polyclonal antibody against S-100 was used, so that S-100 isoforms could not be distinguished (Table 1).
In testis
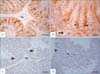
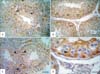
The immunoreaction for S-100 revealed variation between the different studied species. In roosters (Fig. 1A, B), pigeons (Fig. 2A, B) and rabbits (Fig. 2C, D), intense S-100 reaction was seen in Sertoli cells. S-100 was also observed in the endothelial lining of blood vessels in rabbit testis (Fig. 2C). The positively reacted Sertoli cells showed the reaction in their cytoplasm and nuclei. Conversely, no S-100 immunostaining was observed in the Sertoli cells of Sudani ducks (Fig. 1C, D). Moreover, expression of S-100 was absent in different stages of germ cells, Leydig, and myofibroblast cells of the studied species (Figs. 1, 2).
In epididymis
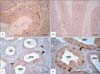
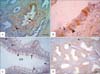
The localization of S-100 had also varied between the different segments of epididymis in the investigated species. In rooster, S-100 specific staining was not detected throughout the whole length of the epididymal duct (Fig. 3A, B). In duck, some basal cells (BC) of epididymal duct showed strong S-100 reaction (Fig. 3C, D). Interestingly, the pigeon DED expressed an alternative pattern of S-100 reactivity between the negative ciliated cells and the distinctly positive non-ciliated cells. Cytoplasm and nuclei of the non-ciliated cells were positive. Some cells in the peritubular connective tissue were also reactive (Fig. 4A, B). Similarly, the efferent ductules (ED) of rabbits showed an alternative pattern of reactivity. However, the distinctly positive cells were the ciliated one (Fig. 4C). These cells showed both cytoplasmic and nuclear S-100 reaction while the non-ciliated cells were completely negative. In epididymis, the nuclei and cytoplasm of BCs exhibited intense S-100 immunostaining (Fig. 4D). Conversely, nuclei and cytoplasm of the principal cells (PCs) were non-reactive. Additionally, the endothelium of blood and lymph vessels as well as some cells in the peritubular connective tissue displayed a weak to moderate reaction along the entire length of the rabbit epididymis (Fig. 4C).
Discussion
S-100 proteins have been implicated in the regulation of cell cycle due to the stimulation of Ndr, a nuclear serine/threonine protein kinase important in the regulation of cell division and cell morphology, in a Ca2+-dependent manner [16]. Additionally, one of the best-characterized functions of S-100 proteins is the regulation of cell morphology, the dynamics of certain cytoskeleton constituents, and the reciprocal relationships of cytoskeleton element via direct and/or indirect interactions with microtubules, intermediate filaments, microfilaments, myosin, and/or tropomyosin [16]. The present study represents the first report that concerned the localization of S-100 in the testis and epididymis of birds and rabbits. In the testis of roosters, pigeons, and rabbits, the S-100 was generally localized in the cytoplasm and nucleus of Sertoli cells. These findings are consistent with numerous approaches (Table 2) that investigated the localization of S-100 in the testis of several mammalian species including bovine [20, 21, 22], sheep [21], rat [21, 24], cat [21, 26], ram, boar, horse, dog [20, 21], buffalo [27], monkey [29], and human [24, 30]. Although the exact testicular function of S-100 is yet unclear, it is striking to note that the bulk of the cell types staining positive for S-100 exhibit special morphological features [24] such as Sertoli cells [20]. S-100 protein in the Sertoli cells is assumed to be involved in the microtubule assembly-disassembly system [29]. Further on, the protein may play a role in the secretory and absorptive functions and may be involved in establishing the blood-testis barrier [21, 27]. Nevertheless, S-100 was not localized to Sertoli cells of the Sudani ducks. Similar findings are identified in European bison [33]. Conversely, S-100 protein is found in Sertoli cells of adult White Peking ducks [31]. The lack of S-100 immunostaining in the testis of Sudani duck is not rational but might be due to species-specific difference. Therefore, morphological studies are necessary to outline the ultrastructure of Sertoli cell in Sudani ducks.
The present work showed that, the rabbit testicular vasculature also exhibited intense S-100 immunostaining. Similarly, endothelial cells of capillaries, veins, and lymphatic vessels are regularly S-100 protein immunoreactive in rat, boar, ruminants, European bison, and human [20, 21, 24, 27, 33]. No expression of S-100 was seen in germ, Leydig, and myofibroblast cells of birds and rabbits. These data are partially concurrent with the previous reports (Table 2). However, Leydig cells are found to be strongly positive for S-100 protein in rat, cat, and human testis and to a lower degree in pig and horse testis [21, 24, 26, 29]. A distinct immunostaining of peritubular cells is only found in the testis of dog, cat, and rat [21, 26].
In the present investigation, S-100 protein was evident in the BCs of epididymal duct in duck, non-ciliated cells of DED in pigeons and the ciliated cells of the ED and BC of the epididymis in rabbits. Additionally, the endothelium of blood and lymph vessels as well as some cells in the peritubular connective tissue was S-100 immunoreactive in the rabbit epididymis. These results are partially consistent with the previous reports of mammals (Table 3). S-100 protein is found in the BCs and PC of epididymis in human [30], bovine [23], donkey [34], buffalo [28], and camel [35]. Localization of S-100 is also seen in the ciliated cells of ED in bovine [23], and in the epididymal vascular endothelium of bison [33], bovine [23], and rat [25]. S-100 is moreover detected in the epithelial cells of epididymal duct in adult White Peking ducks [31]. Collectively, the sites of S-100 localization in the epididymis of poultry and rabbits are partially differed from those in the mammalian epididymis (Tables 1, 3). This might point out to different functional roles. Although a wide variety of diverse functions have been attributed to S-100 proteins, their exact biological role in the epididymis is still not known. S-100 is involved in the absorptive and secretory functions in the intra-testicular excurrent duct system [21, 27]; it may promote similar tasks in the extra-testicular excurrent duct system. S-100 has been associated with several activities including regulation of the diffusion of monovalent cations across membranes and modulation of the physical state of membranes [14]. S-100 may promote a similar task in the excurrent duct system of the male genital tract. S-100 may also participate in the processes of transcytosis and cell contractility [25]. In conclusion, S-100 was localized in the Sertoli cells of roosters, pigeons and rabbits. S-100 was also detected in the BC of ducks, non-ciliated cells of DED in pigeons and ciliated cells of ED and BC in rabbits. Further reports are therefore required to elucidate the actual role of S-100 proteins in male reproduction.



 XML Download
XML Download