

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The role of cerebrospinal fluid (CSF) in maintaining brain function is widely recognized. It effectively absorbs mechanical and thermal stresses encountered by the brain, preventing deleterious consequences. CSF is also considered to have a role in the excretion of potentially toxic byproducts of cerebral metabolism. These functions of CSF can largely be attributed to its fluid state. It was recently suggested that CSF also has a filtration function, allowing movement of water-soluble substances from the brain parenchyma into the CSF. In this manner, solutes entering the brain through the blood-brain barrier, as well as those synthesized by the brain, diffuse freely from the brain interstitial fluid into CSF [1, 2]. There is no barrier isolating the neurons of the brain from CSF; they are freely communicable, and CSF provides them with a stable internal environment. The ionic composition of CSF is extremely important, considering that neurons are excitable cells and very sensitive to electrolyte concentrations in their environment.
It is well established that CSF, including all of its ingredients, is produced by a class of glial cells, called ependymal cells, lining the ventricular surface that is located inside the brain. The choroid plexus, a specialized structure formed by ependymal cells surrounding the blood vessels, plays an essential role in the production and secretion of CSF. However, ordinary ependymal cells that do not form the choroid plexus also participate in the secretion of CSF [3].
Several types of ion channel are responsible for the secretion of electrolytes into CSF. It has been reported that a wide variety of ion channels, including potassium channels, anion channels, receptor operated channels, transient receptor potential (TRP) channels, and an unconventional ion channel, aquaporin 1, are present on the epithelium of the choroid plexus in mammals [3, 4]. The diversity of channels reflects the versatility of the ependymal cells in making up the ionic component of the CSF. Primitive CSF is composed of the amniotic fluid of the embryo after the neuropores have been closed. The neurons are in a close relationship with the CSF from the embryonic period throughout the rest of life as the brain grows and matures during its development. As the neurons of the developing brain proliferate and differentiate, overall tissue metabolism might greatly change, both quantitatively and qualitatively. Therefore, alterations would be expected in the composition of CSF as an environment for the developing neurons, and in the biological characteristics of ependyma that is responsible for neuronal development, in order to adapt to these metabolic transitions. In this regard, it is likely that the expression of the channels in the ependymal cells would also change during the developmental process.
Of those channels expressed in ependymal cells, the functional aspect of the newly discovered TRP channels in the regulation of the ionic composition and chemical sensing of body fluids has been investigated. TRP channels, also called nonspecific cation channels, belong to a superfamily that consists of 6 subfamilies, the members of which are expressed in probably all mammalian organs and cell types [5]. The vanilloid subfamily (TRPV) of TRP has 6 members and is primarily involved in thermal and chemical perception. Liedke et al. [6] detected the m-RNA expression of TRPV4, a member of TRPV, in the epithelial cells of the mouse choroid plexus. TRPV4 was activated by changes in osmotic pressure and temperature [6]. Further research revealed that the TRPV-induced responses, increasing transcellular ion flux as well as paracellular permeability, may allow the epithelial cells to adjust to changes in extracellular osmolarity. Therefore, TRPV4 plays a central role in epithelial homeostasis by modulating epithelial barrier function [7]. TRPV1, TRPV4, and TRPA1, the only member of TRP channel ankyrin subfamily (TRPA), have properties in common. In addition to functioning as thermosensors, they respond to other sti muli, such as mechanical stimuli, and a broad range of chemicals [8-11]. TRPV1, TRPV3, and TRPV4 share a multiligand-binding site for ATP and calmodulin, and thus, these 2 compounds play a key role in generating the different sensitivity and adaptation profiles of these 3 TRPV members [12]. Furthermore, TRPV3 shares 40% identity with TRPV1 [13]. Based upon the observation of TRV4 expression in the epithelium of the choroid plexus, and the functional relationship between TRPV1, TRPV3, TRPV4, and TRPA1, it seems reasonable to assume that these channels are expressed in the choroid plexus.
As mentioned earlier, from a developmental viewpoint, there are 2 reasons why changes in CSF characteristics should be taken into account. First, the osmolarity and ionic composition of primitive CSF formed by the closure of neuropores is different from that of adult CSF, and the overall increase in metabolism caused by the growth of brain tissue possibly affects the characteristics of CSF. The change largely depends on the neuroglia responsible for the production of CSF acquiring functionality through differentiation. The neuroglia may differentiate into ventricular lining ependyma (VLE) or choroid plexus ependyma (CPE), which presumably play different roles in producing CSF. It is not yet known whether the differentiation of these 2 classes of ependyma is associated with the change, or if it is the difference in TRP channel expression pattern during the developmental process.
A classical study using 3H thymidine autoradiography revealed that the formation of VLEs in the rat begins at the fourth ventricle as early as the 13th day of gestation, and at the lateral ventricles on the 16th day [14], while the beginning of choroid plexus morphogenesis was observed on the 14th day [15]. In this period, brain structure changes considerably with the accompanying tissue mass enlargement [16]. Growth of the choroid plexus to the size of that of a newborn is not accomplished in the rat embryo until the 18th day of gestation [15]. Therefore, the essential part of ependymal differentiation into VLE or CPE occurs between the 13th and 19th days of gestation. This study was designed to examine the expression of the selected TRP channels in 2 classes of ependymal cells with different locations, namely VLE and CPE, while they were undergoing development and differentiation. Comparison of the results for the embryonic stage with that for the adult stage may give insight into the adaptation and maturation of ependymal cells.
Materials and Methods
Experimental animals and tissue processing
Female Sprague-Dawley rats weighing 230-250 g were used in this study. A group of females was placed with males overnight. The following morning, vaginal smears were obtained, and animals showing sperm-positive smears were considered to be in day 0 of gestation. On the 13th, 15th, 17th, and 19th days of gestation, groups of pregnant females were anesthetized with ether, sacrificed, and the embryos were collected. Collected embryos were immersed in 10% neutrally buffered formalin (NBF) at 4℃ immediately after extraction from the uterus and kept in this fixative overnight. Fixed embryos were washed, dehydrated, and embedded in paraffin.
For the comparison with expression in adult tissue, 3 adult male rats within the same weight range were used. Under ether anesthesia, the adult rats were perfused transcardially with 10% NBF and stored in NBF overnight. Adult tissue was processed and embedded in paraffin in the same manner as the embryonic tissue.
Serial sections 5-µm thick were prepared, attached to glass slides coated with aminopropyl triethoxysilane, and dried at room temperature. All animal care and procedures were performed according to the guidelines of The Committee for the Care and Use of Laboratory Animals, Kwandong University.
Immunohistochemistry
After deparaffinization and rehydration in a series of ethanol concentrations, the tissue sections were treated for 15 minutes with 3% H2O2 in phosphate buffered saline (PBS 0.1 M, pH 7.4) to block the activity of the endogenous peroxidase. The slices were washed with PBS, followed by incubation in 10% blocking serum (Vector Labs Inc., Burlingame, CA, USA) for 1 hour at room temperature to block nonspecific antigen-antibody reactions. The sections were then incubated with a 1:100 dilution of anti-TRPV1, anti-TRPV3, or anti TRPA1 antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 12 hours at 4℃. In cases where the positive staining reaction was too intense, the primary antibody concentration was decreased 2 to 8 fold.
Next day, the tissues were washed with PBS and treated with biotinylated secondary antibodies (Vector Labs Inc.) diluted in blocking sera for 60 minutes at room temperature. After another 3 washes with PBS, the slices were treated with avidin-biotin complex reagent (Vector Labs Inc.) for 1 hour at room temperature.
For the peroxidase reaction, the tissue sections were washed with PBS and allowed to react for 10 minutes with 0.05% VIP peroxidase substrate (Vector Labs Inc.) solution containing 0.03% hydrogen peroxide. The sections were washed, dehydrated, and coverslips were mounted. Microscopic examination and photography was performed using a BX50 light microscope (Olympus, Tokyo, Japan), and for better visualization, a TE2000 microscope (Nikon, Tokyo, Japan) was used.
Results
The immunoreactivity against TRPV1, TRPV3, and TRPA1 was assessed for 4 regional classes: VLE in the lateral ventricles and the fourth ventricle and CPE of the choroid plexus in the lateral ventricles and the fourth ventricle. The results are summarized in Table 1. A specified level of immunopositivity represents the level expressed in a majority (≥ 60%) of ependymal cells of the class. If 2 levels of expression coexist in an equal ratio, a slash (/) sign is inserted between the specified levels. Five levels are used to denote the intensity of expression, namely no reactivity (-), very weak (-/+), weak (+), moderate (++), and strong reactivity (+++). The positive expression criteria are based on cytoplasmic immunopositivity. A note was added if the nucleus was immunopositive or if there were any other specific findings.
Immunoreactivity of developing VLEs
As the morphogenesis of the brain took place, the cells of ventricular surface showed a variety of patterns of reactivity. In the 13-day embryo, telencephalic vesicles, the third ventricle, and the mesencephalon were clearly identified. Neural epithelium lining the rhombencephalic vesicle was still largely single layered in the dorsal aspect, while that lining the tegmental aspect was organized in multiple layers. There was no clear evidence of choroid plexus formation at this stage of brain development.
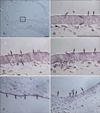
A cluster of columnar neuroepithelial cells lining the medial side of the telencephalic wall showed distinct immunoreativity against all the 3 TRP channel proteins, but at different levels, representing different levels of expression. The regionally clustered neuroepithelial cells showed rather weak reactivity against TRPV1 (1:100). To obtain the same degree of immunopositive intensity, the anti-TRPV3 solution had to be diluted to one half of the TRPV1 antibody concentration. As for the anti-TRPA1 antibody, an 8-fold dilution was necessary. Positive staining reactions were observed in the apical or basal region of the cells, but these did not occupy the entire cytoplasm in this specific region. The nuclei of neuroepithlial cells did not show any expression of the 3 TRP channels (Fig. 1A-D).
Another class showing strongly positive reactivity against these TRP channel proteins was found in the posterior wall of the rhombencephalic vesicle. This region is lined by a single layer of low cuboidal cells with round to oval nuclei. Unlike the TRP-immunopositive cells in the telencephalic wall previously described, the immunopositivity of these neuroepithelial cells was observed in the nuclei as well as in the cytoplasm (Fig. 1E). With the transition from a singlelayered neuroepithelium of the posterior wall to the anterior wall with multiple layers, no immunoreactivity of surface neuroepithelial cells was observed with the increase in the number of cell layers. Some neuroepithelial cells in the deeper layer had reactivity against TRP channel proteins (Fig. 1F).
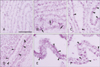
In embryos from the 15th day of gestation, the walls of the neural tube were composed of multiple layers of developing neurons. A horizontal cross section of the fourth ventricle was diamond shaped, with 4 walls having several layers of neuroepithelial cells. VLEs lining the surface of the anterior wall reacted positively to the channel proteins. Moderate reactivity to TRPA1 was localized at the surface of VLEs. The reactivity was weak towards TRPV3 and very weak to TRPV1 (Fig. 2A-C).
Immunoreactivity against TRPA1 was generally higher than that against TRPV1 and TRPV3 from the 15th to 19th days of the development, which was more distinct in the fourth ventricle than in the lateral ventricles. The immunohistochemical staining reaction to antibodies against TRPV1 and TRPV3 did not show a great change in the VLEs lining the lateral and fourth ventricular surfaces from the 17- and 19-day embryos. These cells showed either weak or very weak expression of TRPV1 and TRPV3 during this period (Fig. 2E, G). The reactivity against TRPA1 was moderately positive in the fourth ventricle of the 17-day embryo, while it was weakly positive in the same region of the 19-day embryo and in the lateral ventricles of the 17- and 19-day embryos (Fig. 2F, H).
Within the lateral ventricles of the 15-day embryo, the basal ganglia typically were growing from the anterolateral wall of the telencephalon. VLEs lining the surface of the lateral ventricular cavity exhibited very weak reactivity to antibodies against all 3 channels. Some VLEs lining the region of enlarging basal ganglia showed moderate immunopositivity (Fig. 2D).
During its development, no glycogen compartment of vesicular nature like those observed in the developing CPE was formed in the cytoplasm of the VLE.
Immunoreactivity of developing CPEs
In the brain of the 13-day embryo, no structure considered to be the choroid plexus was found. Primitive choroid plexus composed of cuboidal to columnar epithelium with a mesenchymal core was first observed in the ventricles of the 15-day embryo. The blood vessels residing in the mesenchyme did not appear to be fully developed in this period.
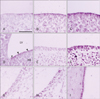
CPEs that formed a covering epithelium were immunostained generally with weak to moderate reactivity against TRPV1, TRPV3, and TRPA1. The overall expression of TRP channels in CPE was greater than that in VLE in our study. Particularly, TRPA1 expression by CPE was greater than TRPV1 or TRPV3 expression in the 15th and 17th day embryos, which decreased in the 19th day in both the lateral and fourth ventricles. A moderate immunopositivity against TRPA1 was mostly visible at the apical region of CPE cytoplasm in the 15-day embryo. The nuclei showed no immunopositivity to the antibodies against all the 3 channels. CPEs sometimes formed an oval vacuolar compartment in the basal region of their cytoplasm at this stage. Some of these compartments exhibited a strong immunoreactive staining reaction against TRPV1 and TRPV3 channel antibodies around their periphery. This feature was observed in the choroid plexus of the lateral ventricles as well as in that of the fourth ventricle (Fig. 3A-C).
The choroid plexus of the 17-day embryo formed a more developed contour. There was multiple branching typical of the choroid plexus, and well-developed blood vessels running inside the stromal mesenchyme beneath the covering CPEs. A moderately positive reaction against TRPA1 was observed in CPE of the 17-day embryo as in the previous stage, while TRPV1 and TRPV3 showed very weak and weak immunopositivity, respectively. Basally located vacuoles found in the previous stage became considerably larger and in the basal portion of the cytoplasm of CPE.
Their size varied a little from cell to cell. Sometimes they were so large that the nucleus and the rest of the cytoplasm were pushed to the small apical region of the cell. These compartments are glycogen accumulations, according to previous studies [17, 18]. Similarly, the 15-day embryo showed a strong positive reaction against TRPV3 which was observed at the periphery of the glycogen and the adjacent cytoplasm. It was the strongest immunopositivity demonstrated in this study. Not all the compartments showed such immunopositivity, and the immunoreactive CPEs tended to be clustered (Fig. 3E). No positive reaction against TRPV1 was observed after this stage.
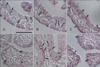
In the 19-day embryo, the lateral and the third ventricles showed a striking shrinkage of the ventricular space, which had not been observed in the previous stages. It was observed that CPEs forming the choroid plexus were in close proximity to VLEs (Fig. 4D). The immunostaining property of the CPE of the 19-day embryo was very similar to that of the VLE: CPEs were very weakly TRPV1-positive and weakly TRPV3- and TRPA1-positive, and showed no difference in reaction intensity between the nucleus and the cytoplasm (Fig. 4A-C).
Glycogen accumulations found in the previous stage increased even more and occupied a much larger area of the cytoplasm in this stage. They were generally uniform in size and hardly showed immunopositivity in the surrounding cytoplasm, except for that glycogen deposit with a weakly TRPV3-positive periphery in the cytoplasm of a few CPEs (Fig. 4B).
Immunoreactivity of the adult ependymal cells
Moderate positive reactions against TRPV3 and TRPA1 were observed in the VLE of all the ventricles (Fig. 2I). The intensity of TRPV1 staining was so weak that it may be considered to be expressed in a very small amount.
CPE in both the lateral ventricles and the fourth ventricle equally expressed moderate immunoreactivity against TRPV3 and TRPA1. The patterns of expression were different: immunopositivity against TRPA1 was found uniformly throughout the cytoplasm, while that against TRPV3 seemed aggregated in a region within the cytoplasm. This allowed the expression of TRPV3 to be seen as a stronger reaction in adult CPE (Fig. 4E, F).
Discussion
As the embryonic brain undergoes its development, VLE and CPE cells show different patterns in the expression of 3 TRP-channel antigens, suggesting different roles in the development of the ventricular system. The developing VLEs generally tend to express TRPA1 more than TRPV1 or TRPV3, and this was observed as early as day 13 in rat embryos. In those embryos, the immunohistochemical reaction was so strong the TRPA1 antibody in the staining solution had to be diluted 8 fold to obtain the same staining intensity. This may mean the level of expression of TRPA1 is 8 times higher than that of TRPV1. Similarly, greater staining of VLE by TRPA1 antibody was consistently observed in both the lateral and fourth ventricles throughout all the stages. Thus, the VLEs of all ventricles express the TRPA1 channel at a higher level than the other 2 channels.
A group of the neuroepithelial cells forming the medial telencephalic wall characteristically expressed all 3 of the channel proteins at a moderate intensity in the 13-day embryos. The expression was detected solely in this limited region of the bilateral telencephalic vesicles. The cells showed the strongest immunopositivity against TRPA1, although reaction against TRPV1 and TRPV3 was also clearly observed when antibody solutions with higher titers were used. These cells were located just above where the interventricular foramen would develop, so these cells might be involved in the formation of the choroid plexus of lateral ventricles during subsequent developmental stages. These cells are not likely to be developing ependymal cells because those are observed in the lateral ventricles at the 16th day of gestation [14]. Since no specific marker has so far been identified with the commencement of choroid plexus morphogenesis, this result means that the TRP channel could be a candidate for a marker for choroid plexus formation in this area.
In the rhombencephalic vesicle, which develops into the fourth ventricle, the posterior wall was composed of a single layer of low cuboidal cells with strong immunopositivity against TRP channels. Because this wall is the portion posterior to the sulcus limitans, it is called the alar plate of the neural tube. Expression of the TRP channels in the alar plate neuroepithelium was observed only in the deeper layer of the anterior part of the telencephalic wall, namely the basal plate of the neural tube where the neuroepithelium consists of several layers of cells. It seems that the expression of TRP channels in cells facing the ventricular space is restricted in the single-layered wall in this region. The reason for this is unknown, but it is hypothesized that it may be associated with the increase in tissue metabolism needed for cell proliferation to produce a wall composed of several layers.
The morphology of the choroid plexus in the adult rat brain was very similar to that in the human brain, taking the form of branching folds and processes located inside the ventricular cavities. Each choroid plexus consists of a single layer of cuboidal to columnar epithelial cells, namely the ependymal cells, situated on a connective tissue core, the stroma. In the connective tissue stroma, there are blood vessels lined with fenestrated endothelial cells. The first evidence of choroid plexus morphogenesis appeared on the 15th day of gestation in our series. Development of the choroid plexus progressed rapidly, reaching the mature contour by the 19th day. The manner of the development of the choroid plexus is consistent with that observed in previous studies [15, 18].
Expression of the 3 TRP channels in the developing CPE is confirmed by the immunohistochemical staining reaction. It appears that the level of expression in CPEs is somewhat stronger than that in VLEs. TRPA1 was shown to express at the highest level, while both TRPV1 and TRPV3 were also expressed in CPE at higher levels than that observed in VLE. Likewise, it has been reported that other membrane channels such as TRPV4 and aquaporin are expressed in the CPE cytoplasm as well as in the plasma membrane [6, 19]. This can be explained in terms of the function of the CPE in trafficking the components of CSF between the blood vessels in the choroid plexus stroma and the lumen of the ventricular cavity. Rougemount and coworkers first suggested that CSF is secreted by the CPE and not by the ultrafiltrate, when they demonstrated that the solute content of nascent CSF differed from that expected for a plasma ultrafiltrate [20]. Recently, it has been generally accepted that the production and secretion of CSF is an active, energy-consuming metabolic process that occurs mainly in the choroid plexus [21]. It is essential for the CPE to carry the ionic components of CSF by way of its cytoplasm without incurring adverse effects. It is important to consider intracellular concentrations when the movement of ions is involved. The main problem with the transcellular route is that it requires that the intracellular osmolarity always be kept between the luminal and basolateral levels [3]. To solve this problem, it is best to carry these ions and electrolytes within transport or pinocytotic vesicles traveling through the CPE cytoplasm. Therefore, it is suggested that these vesicles express the channel protein incorporated into their membrane. Supporting this mechanism, Liddelow et al. [22] recently proposed a pathway involving secreted protein acidic and rich in cysteine and glycophorins, as well as the tubulocisternal endoplasmic reticulum, Golgi complex, vacuoles, lysosomes, and multivesicular bodies.
Another feature shown in this work is that the strongest immunopositivity was found surrounding the cytoplasmic compartment of the CPE in the 15- and 17-day rat embryos. Based on the immunohistochemical reaction, there is a very high level of TRPV1 and TRPV3 expression at the periphery of the oval compartment in the 15-day embryo. This compartment seems to contain the glycogen normally present in the CPE during the development of the choroid plexus. The glycogen compartment, which had been small and located basally in the cytoplasm, became so large that it occupied most of the cytoplasm and was no longer immunoreactive to TRPV1 in the 17-day embryo. In the 19-day embryo, glycogen filled the major portion of almost every CPE cytoplasm of the choroid plexus in both the lateral ventricles and the fourth ventricle, and there was no immunohistochemical reaction against TRPV1 or TRPV3. An electron microscopic study of the development of the choroid plexus showed that the glycogen in the CPE is not surrounded by plasma membrane [17]. In this regard, it is mysterious that the expression of TRPV1 and TRPV3 membrane channels was detected at the periphery of the glycogen. The close relationship between glycogen and TRP is actually apparent in the 15-day embryo only when the glycogen accumulation was small and entirely surrounded by expressed TRP. It is not likely that glycogen deposition is closely associated with the expression of TRP channels. Based on the observation that the majority of glycogen deposits did not show expression of TRP channels at their periphery in the 17-day embryo, the high level of TRPV3 expression located in the basal region of the cytoplasm might coincide with the appearance at this stage of the large glycogen compartment filling most of the basal and central regions of the cytoplasm. As the function of glycogen during the development of the choroid plexus remains unknown, the relationship between glycogen and the TRP channels of the 15-day embryo remains unexplained.
A possible explanation for the existence of glycogen in great measure is the CPE's function in the production and secretion of CSF. Many studies confirm that this is an energy-consuming process involving a variety of channels [3], including TRP [12]. In this instance, glycogen might be a rapidly available energy source. It can be hypothesized that there is an urgent need to change the composition of CSF at this developmental stage because no more glycogen is deposited in the CPE cytoplasm in the following stages, either prenatally or postnatally. If some TRP channels are involved in this process, TRPA1 is most likely to be a candidate, since the expression of this channel appears to persist after the 19th day of gestation. On the other hand, TRPV3 possibly plays a role in the absorption from the blood, for it is expressed in the basal cytoplasm near the blood vessels. In the adult choroid plexus, glycophorins and aquaporin, as well as many ion transporters are expressed in the basal cytoplasm or plasma membrane [3, 19, 22].
CSF is initially derived from amniotic fluid trapped with the closure of the neuropores, and the early amniotic fluid is known to be isotonic with maternal plasma, but gradually becomes hypotonic [23]. Adult CSF is known to be 5 mOsm hypertonic to plasma [24]. Likewise, [Na+] and [HCO3-] of nascent CSF are higher, and [K+] and [Cl-] are lower than that of plasma [25, 26]. Therefore, the differentiation process of CPE and VLE during their development is reflected in the activities of these specialized glia to obtain the required level of ionic composition and osmolarity against the gradients.
In a group of conditions called channelopathies, faulty expression of various channel proteins is observed. Though experimentally induced errors in TRP channel expression have been frequently associated with many consequences in laboratory animals, only a few channelopathies have been identified in which defects in TRP genes in humans are the direct cause of cellular dysfunction, resulting in disease; however, their number is increasing [27]. Much work has to be done to enhance our knowledge of the detailed mechanisms through which TRP channels function. The development and differentiation of the choroid plexus might be a useful subject for research on channelopathies.
The shrinkage of the ventricles was reported by Bayer et al. [16]. They described the shrinkage to start from the 16th day of gestation and continuing to the 19th day in rat. In our studies, the shrinkage was most prominent in the lateral and third ventricles of the 19-day embryo, which is consistent with the results of previous report. This shrinkage was largely due to neuronal migration during the development of basal ganglia, basal telencephalon, and diencephalon [28]. In our work, it also appeared that a decrease in TRP channel expression in the CPE of the lateral ventricles accompanied the shrinkage in this period. It can be hypothesized that the expression of channels in the CPE may be downregulated with the shrinkage of the ventricles. Coinciding with this, there was widespread expression of TRP channels, especially TRPV3 and TRPA1, in the VLE covering the surface of enlarging basal ganglia on the 15th day, implying a possible relationship between the need to make space for the growing brain structure and the expression of channels in the VLE. Some researchers showed that if the brain of chick embryos is intubated in situ with a fine capillary early in its development so that CSF drains out of the ventricular system, then the brain loses its characteristic shape, and cell layers pile on each other in a disorganized fashion, and the cell division continues, but at a slower rate. This consequence was ascribed to the absence of increasing pressure within the ventricular system [29]. This work was criticized in that an additional factor might be the exposure of the ventricular zone cells to proteins from the general extracellular environment of the embryo as a result of puncturing the ventricular system [18].
In conclusion, to investigate the involvement of TRP channels in the modification of embryonic CSF, we used an immunohistochemical method to study their expression in ependymal cells of the ventricular lining and choroid plexus at discrete stages of development. Our findings showed that the TRPA1 expression was observed from the 13th day through the 17th day in VLE as well as CPE and its expression decreased at the 19th day of gestation which appeared to be expressed again in the adult stage. TRPV1 and TRPV3, expressed at a high level in early stage with TRPA1, are likely to function actively in CPE from the 15th through the 17th day when there was a distinct intracellular development and enlargement of the glycogen compartments.

 XML Download
XML Download