

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Although elastic fibers are a well-known component of human fibrous tissues, their distribution in human fetuses has not been examined extensively, possibly because of the identification of these fibers is difficult. In the adult human body, elastic fibers generally coexist with smooth muscles, and both show a specific organizational relationship [1-3]. If fetal elastic fibers also coexist with smooth muscles in the head region, then their distribution would be very limited, for example, their distribution in arterial walls and some cartilages, because smooth muscle is fairly scant in the fetal head. We are aware of limited examples of smooth muscle that are not closely related to vessels in the fetal head: the orbitalis muscle behind the orbit [4] and the ciliary muscle. However, elastic fibers can be present without associated smooth muscle in specific structures such as elastic cartilages, the nuchal ligament [5], the yellow ligament (ligamentum flavum) [6, 7], the skin dermis [8], and the facet capsule of the vertebral column [9]. In the last case, Shiraishi et al. [9] used human fetal material for their research. Likewise, specific fascial structures in the human pelvic floor contain elastic fibers without any association with smooth muscle [10-13]. In contrast, information about elastic fibers in the head connective tissue is limited: only a few reported studies of experimental animals [14-16] and humans [17] have demonstrated that the temporomandibular joint contains elastic fibers.
A notable constituent of normal elastic fibers is osteopontin, an acidic matrix protein that is expressed mainly in mineralized tissues [18]; therefore, elastic fibers facilitate calcification [19]. In fact, in the ligamentum flavum, changes in elastic fiber configuration have been well described in relation to calcification [6, 7]. We have noted that Meckel's and Reichert's cartilages in fetal heads undergo significant changes in morphology to form the final derivatives [20-25]. Calcification can occur in Meckel's and Reichert's cartilages at specific sites such as ear ossicles. Therefore, we hypothesized that these cartilages are likely to accompany elastic fibers as an intermediate developmental step before attaining their final morphology. Consequently, the aim of this study was to examine the distribution of elastic fibers in late-stage human fetuses. To identify the elastic fibers, we employed both aldehyde-fuchsin and elastica-Masson staining.
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). We examined paraffin-embedded sagittal sections of 15 late-stage fetuses (28-37 weeks of gestation; crown-rump length, 220-320 mm). All the specimens were part of a large collection kept at the Embryology Institute of the Universidad Complutense, Madrid, and were the products of urgent abortion, miscarriages, or ectopic pregnancies that were managed at the Department of Obstetrics of the University. The donated fetuses were fixed and stored for >3 months in 10% v/v formalin solution. After trimming the tissue mass, decalcification was performed for 2-5 days at room temperature using Plank-Rychlo solution (7% [w/v] AlCl2 · 6H2O, 3.6% HCl, 4.6% HCOOH; WAKO, Tokyo, Japan). After routine procedures for paraffin-embedded histology, 10-µm thick sections were prepared at intervals of 0.5-1 mm. To identify elastic fibers, we chose 2 different types of staining: 1) aldehyde-fuchsin staining [26, 27] and 2) elastica-Masson staining, a variation of Masson-Goldner staining [28-30]. The aldehyde-fuchsin staining was performed after overnight post-fixation of the section on the glass slide with Bouin's fixative. Because of the severe conditions used for decalcification, immunohistochemistry was not possible with these specimens (data not shown).
Results
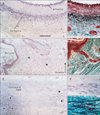
As positive controls for aldehyde-fuchsin and elastica-Masson staining, we used the elastic laminae of the arterial walls, nuchal skin, and elastic cartilage of the external auditory meatus (Fig. 1). The nuchal ligament had not yet developed in these specimens. The elastic laminae of the external carotid artery were selectively stained and colored bright violet using aldehyde-fuchsin (Fig. 1A) or black using elastica-Masson (Fig. 1B). The nuchal skin dermis contained elastic fibers, which were identified as violet or black dots (Fig. 1C, D). We did not compare the direction of these fibers with that of other parts of the skin. The cartilages of the external auditory meatus, which were identified as irregularly shaped fragments, contained elastic fibers in the center of each fragment (Fig. 1E). The ligament-like structure between the cartilage fragments was also composed of elastic fibers (Fig. 1F, G). The morphology of the elastic fibers stained by these 2 methods did not differ critically. In addition, parotid gland acini and osteocytes in areas of endochondral ossification were darkly stained with aldehyde-fuchsin. Such clear discrimination of elastic fibers from other fibrous tissues by aldehyde-fuchsin staining was evident in 11 of the 15 specimens examined, while elastica-Masson staining consistently demonstrated elastic fibers in the arterial walls, skin dermis, and external ear cartilage. Therefore, most of the descriptions below are based on observations of these 11 specimens, for which both types of staining were possible.
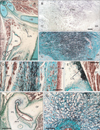
The connective tissue around the maxilla and mandible, that is, the sphenomandibular ligament consistently showed elastic fibers (11/11), but they were restricted to the anterior layer of the superior half of the ligament (Fig. 2). Most of the fibers ran along the superoinferior axis (Fig. 2B, C). The temporomandibular joint disk also contained irregularly arrayed elastic fibers at the superolateral end (Fig. 2D). Although they were near or adjacent to the sphenomandibular ligament, we did not observe elastic fibers in the discomalleolar ligament. In the fascia covering striated muscles in the head, a fascia of the stylopharyngeus muscle specifically contained abundant elastic fibers (Fig. 2E-G); this fascia corresponded to the anterior margin of a large fatty tissue mass forming the prestyloid compartment of the parapharyngeal space or the prestyloid space [31, 32]. However, elastic fibers were found along the tensor veli palatini muscle or the primitive styloid process or Reichert's cartilage. The upper extension of the carotid sheath, as well as the prevertebral lamina of the deep cervical fascia, did not contain elastic fibers. In 3 of the 15 specimens, Meckel's cartilage contained elastic fibers in the anterior part of the perichondrium, but they were not stained with aldehydefuchsin (Fig. 2H-J). In addition, the periosteum of the mandible sometimes (3/11) contained elastic fibers at the lateral aspect facing the masseter muscle (Fig. 5C). The nasal mucosa contained a few elastic fibers along the periosteum of the hard palate (Fig. 5B).
We observed elastic fiber accumulation in ligament- or fascia-like structures that connected the hyoid bone, thyroid cartilage, and cricoid cartilage. Among these structures, the lateral thyrohyoid ligament (Fig. 3A-C) and the cricothyroid ligament (Fig. 3E) were almost purely composed of irregularly arrayed elastic fibers. The former connected the greater horn of the hyoid bone to the superior horn of the thyroid cartilage, while the latter connected the thyroid and cricoid cartilages along and near the anterior midline. These cartilages and the bones themselves were accompanied much or less elastic fibers in the perichondrium or periosteum. The epiglottic cartilage also contained elastic fibers, but they were interrupted by glands that invaded the cartilage. The vocal fold had not developed in the examined specimens. The submucosal tissue of the pharynx and larynx also contained a few elastic fibers. However, the laryngeal and pharyngeal wall muscles usually had no accompanying elastic fibers with 1 exception: abundant elastic fibers ran along the superior-inferior axis in a space between the palatopharyngeus muscle and the posterior pharyngeal mucosa (Fig. 3D). The submucosal tissue contained elastic fibers around the oral cavity, especially at the lateral angle near the opening of the parotid duct. The associated cartilage contained elastic fibers at and around the pharyngotympanic tube, but they were restricted to the junction between the lateral and medial laminae of the cartilage (Fig. 3F, G). The teeth were unaccompanied by elastic fibers.
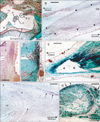
Notably, elastic fibers had accumulated in the middle ear at 3 specific sites: 1) in the tendon of the stapedius muscle (Fig. 4A, B, E), 2) in the capsules of articulations between the malleus and the incus and between the incus and the stapes (Fig. 4C, D), and 3) in an interface area between the tensor tympani muscle and its tendon (Fig. 4F, G). Among these 3 sites, the latter 2 showed a constant morphology in all 11 examined specimens, which was in contrast with the observations of the joint capsules (3 of 11 specimens). However, elastic fibers in the joint capsule appeared to insert into the cartilage covering the ear ossicles. In the middle ear, the amount of elastic fibers was greatest in the tendon of the stapedius muscle. The epimysium of the middle ear muscles, the periosteum of the ear ossicles, and the tympanic membrane did not contain any elastic fibers. We failed to observe the base of the stapes attaching to the fenestra vestibuli. In addition, no elastic fibers were observed in the internal ear or eyeball.
In our observations, we did not find any clear differences between stages: individual differences in the strength of staining, especially with aldehyde-fuchsin appearing to mask any differences between stages. Because of our extensive trimming of the specimens, the face and eyelid could not be included. The orbitalis muscle behind the orbit was unaccompanied by elastic fibers (Fig. 5A). These results are summarized in Table 1.
Discussion
This study revealed the distribution of elastic fibers in the fetal head and neck. It seems reasonable for elastic fibers to be abundant in ligament- or fascia-like structures in the larynx and pharynx because of functional demands during adulthood. At the same time, during development, these bands of elastic fibers appeared to maintain the topographical relationship between cartilages. Similar examples were seen in the external ear cartilages and the pharyngotympanic tube cartilage. In the former, cartilage fragments in the external meatus were united by elastic fiber bands as the meatus expanded. In the latter, the elastic fibers appeared to unite and fix the lateral lamina of the tubal cartilage to the medial lamina against traction by the tensor veli palatini muscle. The finding that the orbitalis muscle (the largest mass of smooth muscle in the fetus) contained no elastic fibers was unexpected; the muscle appeared to change into collagenous tissue through a process that differed from that of the sphenomandibular ligament (see below). Although there is little information about the distribution of elastic fibers in adults, we found that, in fetuses, 1) a mini-version of elastic fiber distribution compared to that in adults and 2) a different specific developmental pattern of connective tissues.
In fetal heads, Meckel's and Reichert's cartilages show a significant change in morphology as the final derivatives form [22-25]. Calcification can occur in either of these cartilages at specific sites such as the ear ossicles and styloid process. Changes in the histology of Meckel's cartilage have been well described in late-stage fetal and postnatal life [20, 21], but elastic fibers have received scant attention. Elastic fibers facilitate calcification because they accompany several proteins such as osteopontin that play a key role in bone and cartilage development as mentioned previously. Accordingly, these cartilages are likely to accompany elastic fibers in the intermediate developmental step before they attain their final morphology. Likewise, in the ligaments and fascia of the fetal head, the elastic fiber content is also likely to reflect an intermediate morphology between the cartilage and final collagen-dominant structure because elastic fibers are an important component of the scaffolds on which tissue development and remodeling depend [33, 34]. However, there has been no information about the elastic fiber contents of the adult sphenomandibular ligament and the posterior marginal fascia of the prestyloid space. The former ligament is a remnant of Meckel's cartilage [22], whereas we have hypothesized that the latter fascia is likely to be one of the derivatives of Reichert's cartilage (submitted as another paper). Shin et al. [35] termed the latter fascia "the tensor-vascular-styloid fascia." In contrast with the sphenomandibular ligament, we did not find elastic fibers in the adjacent discomalleolar ligament. However, if the elastic fibers represent an intermediate morphology during the differentiation of Meckel's cartilage, a further study seems necessary to identify elastin (not elastic fibers) in materials at a stage earlier than the present specimens.
The amount of elastin in tendons is usually limited to 0-2% of the dry mass, in contrast to 65-80% for collagens [36], with a few exceptions such as the vincular membrane in the phalanges of the hand [37] and tail tendons of animals [38]. The limited amount of elastin is thought to contribute to the recovery of wavy collagenous fibers after tendon stretching [36]. Likewise, ligaments contain few elastic fibers, with some exceptions such as the yellow ligament and nuchal ligament of the vertebral column [5, 39]. Notably, in this study, 2 striated muscles in the middle ear accompanied abundant elastic fibers at the muscle-tendon interface as well as in the tendon itself. Although striated muscle fibers have their own elasticity [40], the covering fascia of skeletal muscle contains various amounts of elastic fibers [41]. However, rather than elastic fibers, it is thought that the elasticity of the fascia is provided by overlaid collagenous meshworks, each displaying a different orientation. Thus, in contrast to smooth muscle (see Introduction), elastic fibers do not seem to play a critical role in the function of striated muscle, with a few exceptions such as the levator ani muscle at the pelvic floor [10, 42, 43]. We found that the middle ear muscles did not carry elastic fibers in either the epimysium or the covering fascia; this was not observed in the pelvic floor muscle. Under the specific conditions of constant vibration (the so-called acoustic oscillation) from the tympanic membrane, it seems reasonable that the elastic fibers in the tendons and joint capsules would reduce mechanical stress on the ear ossicles. The tendons attached to the ear ossicles develop through a complex process involving the regression of the pharyngeal arches [23, 25]. Thus, as hypothesized for elastic fibers in the aforementioned ligament and fascia, we cannot rule out the possibility that the elastic fiber content observed in the middle ear represents an intermediate morphology specific to late-stage fetuses.
Overall, the following were observed in human fetuses: 1) a mini-version of elastic fiber distribution compared to that in adults and 2) a different specific developmental pattern of connective tissues. Guadall et al. [44] reported that, during hypoxia, fibulin (a major component of microfilaments accompanying elastic fibers) is upregulated through a hypoxia-inducible factor 1-dependent mechanism. Thus, the latter morphology that was specific to fetuses might be the result of adaptation to hypoxic conditions during development. This study was limited in that the 2 types of staining that we employed were insufficient for identifying elastin or immature elastic fibers such as oxytalan and elaunin [45]. Because of the severe conditions used for decalcification, the present materials were unsuitable for elastin immunohistochemistry. When con sidering the intermediate morphology of the pharyngeal arch cartilages (see above), the immature status of the elastic fibers should be considered.



 XML Download
XML Download