

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Severe ascorbic acid-insufficiency (scurvy) results in defects in collagen synthesis [1], and is associated with connective tissue lesions such as vascular purpura, bleeding and gum abnormalities [2]. Given that organic bone matrix is composed mainly of type I collagen [3], ascorbic acid-insufficiency has been speculated to play a role in bone abnormalities. For example, inhibition of new bone formation after bone grafting, and a delay in bone regeneration after injury, have been observed in scorbutic guinea pigs [4, 5]. Moreover, in humans, ascorbic acid-insufficiency causes bone abnormalities such as microfracture, osteolysis and osteoporosis [6-8]. As a corollary, epidemiologic studies have shown that supplementation with ascorbic acid may enhance bone formation and overall bone health in humans [9, 10].
The synthesis of ascorbic acid from glucose in most animals negates the need for dietary intake of ascorbic acid by food. However, due to the lack of L-gulono-γ-lactone oxidase, an enzyme essential for the synthesis of ascorbic acid which catalyzes the conversion of L-gulono-γ-lactone to L-ascorbic acid [11-13], humans must obtain ascorbic acid in their diet. The ability of most experimental animals to synthesize ascorbic acid is a major impediment to the study of the in vivo effect of ascorbic acid in animals. Accordingly Gulo-/- mice, which contain a defect in the L-gulono-γ-lactone oxidase gene and are unable to synthesize ascorbic acid [14], are considered a useful model for the investigation of the in vivo effects of its supplementation. For example, we recently demonstrated the preventive effect of vitamin C on the development of acute hepatic inflammation in Gulo-/- mice [15].
During the course of a previous investigation of the general characteristics of Gulo-/- mice [16], skeletal changes, including chondrocostal junction thickening and multiple fractures, were clearly evident after 5 weeks withdrawal of ascorbic acid. To determine the specific mechanisms involved in these lesions, we investigated the effect of ascorbic acid insufficiency on bone metabolism in Gulo-/- mice.
Materials and Methods
Mice
Gulo+/- breeding pairs were obtained from the Mutant Mouse Regional Resource Center, MMRRC (University of California, Davis, CA, USA). We determined the genotypes of the offspring by polymerase chain reaction as recommended in the literature [14]. Gulo-/- and C57BL/6 wild type (WT) mice were maintained in specific pathogen free conditions in the animal facility at the Seoul National University College of Medicine. Twelve-week-old male mice were used in this experiment, and the protocol was reviewed and approved by Ethics Committee of the Seoul National University.
Supplementation and withdrawal of ascorbic acid
Gulo-/- mice were supplemented with ascorbic acid (3.3 g/l, Sigma, St. Louis, MO, USA) in water to maintain the general health of the mice for 12 weeks. The mice were divided into three groups. One group received continuous ascorbic acid supplementation until the end of the experiment, referred to as "ascorbic acid-sufficient Gulo-/- mice." The second group underwent complete withdrawal of ascorbic acid supplementation for the last 3 weeks of the experiment, referred to as "3-week ascorbic acid-insufficient Gulo-/- mice." The third group underwent a withdrawal of ascorbic acid supplementation for the last 4 weeks of the experiment, referred to as "4-week ascorbic acid-insufficient Gulo-/- mice." To determine the general physiological changes in Gulo-/- mice in response to ascorbic acid insufficiency, the body weight of all mice was measured at the beginning and end of the experiment.
Biochemical analysis
At the conclusion of the experiment, blood was collected from the intra-orbital plexus with a heparinized capillary tube. Plasma was obtained from each blood sample by centrifugation at 14,000 rpm for 30 minutes. The plasma levels of calcium, phosphorus, and alkaline phosphatase (ALP) were measured with a chemistry analyzer (Hitachi 7070, Hitachi Science Systems, Ltd., Hitachinaka-shi, Japan). The plasma level of osteocalcin was measured with a mouse-specific enzyme-linked immunosorbent assay kit following the manufacturer's protocol (Biomedical Technologies, Inc., Stoughton, MA, USA).
Determination of tibia dry weight
Mice were sacrificed by cervical dislocation under ether anesthesia. The left leg was dissected and the surrounding soft tissues were removed. After de-fatting with a mixed solution of chloroform/ethanol (2:1) for 24 hours, the tibia was flushed out with saline to remove the remnant marrow element. The fat free tibia was then dried in an oven at 80℃ over 48 hours. The tibia dry weight was measured with a microbalance (AEX-200G, Shimadzu Co., Kyoto, Japan).
Histologic examination
The dissected right tibia was fixed in 4% paraformaldehyde for 24 hours after removal of the surrounding soft tissues. Decalcification was performed with a 10% ehylenediaminetetraacetic acid solution for 5 days [17]. The fixed and decalcified tibia was embedded in paraffin and sectioned at 4 µm. Paraffin bone sections were stained with hematoxylin and eosin (Sigma) according to the manufacturer's instruction.
Statistical analysis
Results are presented as the mean±SD. Differences among the groups were tested using the Kruskal-Wallis test, and subsequent comparisons of each group were performed using a Mann-Whitney test. Data were analyzed using PASW for Windows ver. 18.0 (SPSS Inc., Chicago, IL, USA). All statistical outcomes were based on a two-sided test, and a P<0.05 was regarded as statistically significant.
Results
Decreased weight of Gulo-/- mice after 4 weeks of ascorbic acid withdrawal
No significant differences in the body weight between mice in any of the four groups were observed prior to the experimental period (Fig. 1A). Drinking water was replaced with water that did not contain ascorbic acid at the indicated times for each of the ascorbic acid-insufficient Gulo-/- groups in the Materials and Methods. We observed a significant decrease in body weight in the 4 week ascorbic acid-insufficient Gulo-/- mice. These data indicate that an extended period of ascorbic acid insufficiency had a negative weight on the weight of the animals.
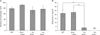
Decreased plasma osteocalcin levels in Gulo-/- mice in response to ascorbic acid withdrawal
We hypothesized that the loss in body weight of the Gulo-/- mice in response to ascorbic acid withdrawal might be related to defects in bone metabolism. To test this hypothesis, we measured the plasma levels of calcium, phosphorus, ALP and osteocalcin in mice in each of the experimental groups at the termination of the experiment. Plasma levels of calcium (Fig. 2A) and phosphorus (Fig. 2B) did not exhibit any significant differences among the groups. The plasma level of ALP, a known non-specific marker of bone formation, was higher in ascorbic acid-sufficient Gulo-/- mice than in mice in the other groups, although this failed to reach statistical significance (Fig. 3A). However, plasma levels of osteocalcin, a specific marker of bone formation, were significantly lower in both 3- and 4-week ascorbic acid-insufficient Gulo-/- mice, compared to WT and ascorbic acid-sufficient Gulo-/- mice (Fig. 3B).
Effect of ascorbic acid insufficiency on tibia dry weight in Gulo-/- mice
To examine the net effect of ascorbic acid-insufficiency on bone metabolism, tibia dry weight was measured at the conclusion of the experiment (Fig. 4A). The average tibia dry weight in ascorbic acid-sufficient Gulo-/- mice was significantly higher than that in the other three groups. No significant differences were among the WT, 3-week and 4-week ascorbic acid-insufficient Gulo-/- mice with respect to dry tibia weight. To adjust for the effect of general growth difference, we compared the ratio of the tibia weight to final body weight between the WT and ascorbic acid-sufficient Gulo-/- mice. This ratio was significantly higher in the ascorbic acid-sufficient Gulo-/- mice compared to WT mice (Fig. 4B).
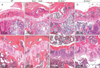
Effect of ascorbic acid insufficiency on trabecular bone architecture in Gulo-/- mice
To evaluate alterations in bone architecture in response to ascorbic acid withdrawal, we next carried out histological examination of mice in the four groups. We observed the growth plate of the tibia in all four groups, indicating that bony growth was not complete at that age (Fig. 5). In both WT and ascorbic acid-sufficient Gulo-/- mice, metaphyseal trabecular bone was well organized (Fig. 5A, B) and trabecular bone was attached to the growth plate (Fig. 5E, F). However, in both the 3- and 4-week ascorbic acid-insufficient Gulo-/- mice, trabecular bone volume was significantly decreased near to the growth plate (Fig. 5E, F), and there was a significant decrease in trabecular bone attachment to the growth plate (Fig. 5G, H). These findings indicate a derangement in new bone formation in response to ascorbic acid insufficiency.
Discussion
Ascorbic acid is a well-known co-factor for collagen synthesis, and a variety of abnormalities in connective tissues, such as scurvy and chondrocostal fracture, occur under conditions of ascorbic acid insufficiency [1, 18]. The extensive physiological changes that occur in humans in response to ascorbic acid insufficiency, including decreased total cholesterol, high density lipoproteins and catecholamine levels, and increased low density lipoproteins and weight loss [14], are recapitulated in Gulo-/- mice. Several reports point to the role of ascorbic acid in the generation and maintenance of bone in animals. For example, the osteogenic disorder Shionogi rat is unable to synthesize ascorbic acid due to the lack of L-gulono-γ-lactone oxidase, reflecting the situation in humans and Gulo-/- mice. The accumulation of malformed collagen inside the osteoblast rough endoplasmic reticulum in these rats in response to conditions of dietary ascorbic acid insufficiency, has been shown to result in detachment of osteoblasts [19]. In addition, reductions in bone hydroxyproline content and osteocalcin mRNA expression [20, 21], as well as in bone and serum ALP activity [21], have been reported in ascorbic acid-insufficient guinea pigs. Moreover, spontaneous fractures and severe impairment of osteoblast differentiation are characteristic of sfx mice, which cannot synthesize ascorbic acid due to deletion of the L-gulono-γ-lactone oxidase gene [22]. Collectively, these reports indicate that ascorbic acid is an important factor for maintaining the structure and function of bone, although the factors and mechanisms involved in this process have to date been largely unknown. Accordingly, we set out in this study to characterize the consequences for bone metabolism of ascorbic acid insufficiency in Gulo-/- mice.
Four weeks of ascorbic acid insufficiency resulted in a decrease in body weight in Gulo-/- mice (Fig. 1). However, there was no evident weight change in Gulo-/- mice after 3 weeks of ascorbic acid withdrawal. This result is in agreement with our previous report on time- and organ-specific changes in in vivo ascorbic acid concentration in Gulo-/- mice [16]. Interestingly, despite the absence of weight loss, there was a remarkable loss of trabecular bone volume near to the growth plate in Gulo-/- mice after 3 weeks of ascorbic acid withdrawal (Fig. 5). Given that similar results have been reported in ascorbic acid-deficient guinea pigs [21, 23], it appears that ascorbic acid-insufficiency exerts its primary effect on bone metabolism.
ALP and osteocalcin are representative markers of bone formation [24]. Because plasma levels of both ALP and osteocalcin are increased by activation of osteoblasts, they are taken to directly reflect changes in bone formation in vivo [24, 25]. We observed significant decreases in plasma levels of osteocalcin, but not ALP, in Gulo-/- mice in both the 3- and 4-week ascorbic acid insufficiency groups (Fig. 3A, B). This is conceivably due to the presence in the serum of these animals of an isoform of ALP expressed in the liver, which is one of the major sources of blood ALP [26]. Because ascorbic acid plays an important role in the production of a variety of essential enzymes, especially in the liver, continuous supplementation with ascorbic acid may result in increased ALP levels in mice in the ascorbic acid-supplemented Gulo-/- group. While we observed no differences in serum calcium or phosphorus levels between any of the mouse groups (Fig. 2), osteocalcin production was decreased and bone volume was reduced in ascorbic acid-insufficient Gulo-/- mice. It appears, therefore, that bone formation by osteoblasts was impaired in ascorbic acid-insufficient Gulo-/- mice. Taken together, these data indicate that the abnormalities in bone formation in ascorbic acid-insufficient Gulo-/- mice are related to a decrease in osteocalcin levels, although the exact mechanism involved requires further investigation.
Both tibia weight, and the ratio of the gross weight of the tibia to total body weight, were higher in ascorbic acid-sufficient Gulo-/- than in WT and ascorbic acid-insufficient Gulo-/- mice (Fig. 4A, B). Moreover, although it was not statistically significant, there was a trend towards an increase in plasma osteocalcin level in the Gulo-/- ascorbic acid-sufficient mice. While the concentration of ascorbic acid in the drinking water (3.3 g/l) is higher than that intrinsically produced by mice, these data indicate that sufficient- or mega-dose uptake of ascorbic acid could enhance bone formation and maintain bone health.
Epidemiological reports of a beneficial effect of mega-doses of ascorbic acid on bone health include a positive association between ascorbic acid intake and bone mineral density (BMD) of the spine and hip in postmenopausal estrogen/progestin trials [9]. Moreover, it has been reported that elevated total ascorbic acid intake is associated with a reduction of femoral neck and trochanter BMD loss in men with low calcium or low vitamin E intake [10]. In addition, femoral neck BMD in male nonsmokers has been shown to be positively correlated with total ascorbic acid intake [10]. As a caveat, however, other studies have failed to find a positive correlation between ascorbic acid and bone health [27, 28], suggesting that further studies on this association are warranted.
Well-organized trabecular bone was observed in WT mice and ascorbic acid-sufficient Gulo-/- mice (Fig. 5A, B). In addition, the attachment of trabecular bone to the growth plate in these animals was intact (Fig. 5E, F). In contrast, in ascorbic acid-insufficient Gulo-/- mice, we observed striking reductions of both trabecular bone volume in the proximal metaphysis near the growth plate, and trabecular bone attachment to the growth plate (Fig. 5C, D, G, H). Because the area just distal of the growth plate is the most active site for new bone formation, we speculate that this is the reason that the reduction in the trabecular bone was more noticeable in the proximal metaphysis near the growth plate than at any other site. On the basis of the previous reports of trabecular bone reduction in ascorbic acid insufficient animal models [18, 23, 29], we anticipated that other bony abnormalities such as cortical bone thinning, growth plate reduction, and malalignment would occur after a more prolonged period of ascorbic acid withdrawal than was evaluated in this experiment.

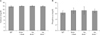
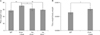
 XML Download
XML Download