

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Skeletal muscles are each connected to a tendon, which in turn is often joined to a ligament. Thus, discrimination between the tendon and the ligament is sometimes difficult, as in the case of the patellar tendon and ligament. A skeletal muscle sometimes originates from or inserts into a ligament: typical examples are seen around the elbow and knee joints. Thus, parts of the supinator muscle originate from the annular ligament of the radius (ANL), and both muscle and ligament take a winding course. The sacrospinous ligament (SSL) has been illustrated as a triangular structure with a medial base and a lateral apex [1, 2]. Hammer et al. [3] defined the SSL as a contorted frustum comprising a single cone with elliptic planes. Henle [4] and Fick [1] described the SSL as being interspersed by the coccygeus muscle fibers, and Hammer et al. [3] confirmed this fact by using the plastination technique. Both the "supinator and ANL" and the "coccygeus and SSL" combinations share a common feature in that the directions of their fibers are very similar in adults.
Mechanical loading by skeletal muscles is required for tendon regeneration and repair [5, 6]. Likewise, dynamic loading by skeletal muscles is critically important for the morphogenesis of tendon and bone. During fetal development, early muscle contraction confers a specific morphology on a tendon [5, 7], and in turn, the tendon confers a specific shape on the bone [8, 9]. Does the inductive relationship between a muscle and a tendon also apply to a muscle and a ligament? In the present study, by focusing on two muscle and ligament pairs, we investigated the possibility that the two structures might show coordinated development and growth, i.e., that the muscle confers a suitable morphology on the ligament. Nevertheless, during dissection of adult cadavers for our educational curriculum, we had an impression that these ligaments grow in combination with degeneration of parts of the muscles because the fiber directions were almost the same and because the muscles sometimes developed poorly.
Development of the ANL during the fetal stage has been described by many research groups [10-13], but its close relationship to the supinator has been referred to in only a single sentence: "The initial ANL is strongest posteriorly at the origin of the supinator muscle" [14]. Does the supinator develop prior to the ANL around the head of the radius? Fetal development of the coccygeus muscle has been described by several groups [15-17]. Niikura et al. [16] stated that the initial SSL is formed by a thick aponeurosis between the sacrococcygeus posterior muscle and the dorsolateral surface of the coccygeus muscle. Does the SSL develop from an intermuscular fascial structure? Consequently, to consider the possibility of coordinated development and growth between muscle and ligament, the aim of this study was to clarify the development of these two muscle and ligament pairs in human fetuses.
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh, 2000). We examined the histology of paraffin-embedded specimens of 1) 15 mid-term human fetuses (crown-rump length [CRL], 55-165 mm; approximately 10-22 weeks of gestation) for observations of the supinator muscle and ANL and 2) 10 larger human fetuses (CRL, 150-260 mm; approximately 20-32 weeks of gestation) for observations of the coccygeus muscle and SSL. All the fetuses were part of the large collection kept at the Embryology Institute of the Universidad Complutense, Madrid, being the products of urgent abortion, miscarriages, or ectopic pregnancies managed at the Department of Obstetrics of the University. Approval for the study was granted by the ethics committee of the university.
The donated fetuses were fixed with 10% v/v formalin solution for more than 3 months. After division into the head and neck, thorax, upper extremities, abdomen, and pelvis with part of the thigh, and the other parts of lower extremities, the 15 specimens for the study of the elbow were decalcified by incubating them at 4℃ in a 0.5 mol/l EDTA solution (pH 7.5; Wako, Tokyo, Japan), while the other 10 larger specimens were decalcified with 5% v/v nitric acid at room temperature. Routine procedures for paraffin-embedded histology were performed. Five- (or 10-) µm-thick sections of the elbow (or pelvis) were prepared at intervals of 10-20 (100-200) µm. All elbows were cut transversely across the long axis of the radius, while for the pelvis, horizontal (3 specimens) or sagittal (7 specimens) sections were prepared. Most of the sections were stained with hematoxylin and eosin, Masson trichrome, or silver staining, while some sections of the smaller fetuses were studied via immunohistochemisty with mouse monoclonal anti-human desmin (1:50, M0760, Dako, Glostrup, Denmark) to show the striated muscle fibers. Autoclave pretreatment was not performed because of the loose nature of the fetal tissues. The secondary antibody (Dako Chem Mate Envision Kit, Dako) was labeled with horseradish peroxidase (HRP), and antigen-antibody reactions were detected via an HRP-catalyzed reaction with diaminobenzidine, followed by counterstaining with hematoxylin.
Results
Supinator muscle and the ANL
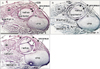
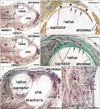
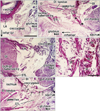
At 10 weeks, although it was still composed of loose fibrous tissue, the ANL was already evident in the deep side of the anconeus (Fig. 1). The supinator muscle originated from a ridge of the cartilagenous ulna via a long distinct tendon (Fig. 1A, B). Notably, at a more proximal level, the supinator tendon became thicker and was continuous with the ANL. In the superficial side of the tendon and ligament, an intermuscular fascia extended between the extensor carpi radialis and anconeus muscles: this fascia was much thinner than the ANL. However, at 12-15 weeks, the intermuscular fascia became thick, and at 18 weeks, it was much thicker than the ANL (Fig. 2). Thus, near the origin from the ulna, the supinator muscle fibers were sandwiched by the ANL and the intermuscular fascia or band (Fig. 2B, D). At 18 weeks, the supinator tendon was tightly attached to the ANL and the tendon was difficult to discriminate from the ligament. Parts of the supinator muscle were still attached to the head of the radius. We observed several thick muscle fibers with multiple nuclei in the supinator along the head of the radius (Fig. 2F). At 22 weeks, both the ANL and supinator muscle were well developed. The thick and tight ANL contained several long muscle fibers originating from the supinator muscle (data not shown).
Coccygeus muscle and the sacrospinous ligament
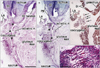
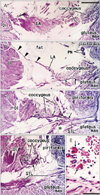
At 18 weeks, the SSL was identified as a loose fibrous band that appeared to be an insertion tendon of the coccygeus muscle (Fig. 3). However, some muscle fibers of the coccygeus were already embedded in the loose ligament (Fig. 3C, D). The primitive ligament was not yet connected with the sacrum. The sacrococcygeus posterior muscle [16] appeared to have already united with the coccygeus. At 28 weeks, the obturator internus muscle was very thick and extended more posteriorly than the tendon curving along the ischium. The tendinous arch of the levator ani muscle was long and loose. An intramuscular tendon was evident in the coccygeus muscle, and this extended outside the muscle along the inferior aspect (Fig. 4). Near the tendon, some muscle fibers of the coccygeus were much thicker than others and had lost their myotube morphology (Fig. 4E). The sacrotuberous ligament was well developed near the ischial tuberosity at 28 weeks, but appeared to connect between the gluteus maximus muscle and the tuberosity (Fig. 4D). At 31 weeks, the intramuscular tendon of the coccygeus muscle was connected with the growing sacrotuberous ligament (Fig. 5). In the posteroinferior margin of the coccygeus muscle, we found a thick bundle of collagenous fibers, which corresponded to the final SSL (Fig. 5D) in view of its topographical relationship with the sacrotuberous ligament. The SSL contained irregularly arrayed muscle fibers (Fig. 5E).
Discussion
Many ligaments in the human body do not apparently develop in association with muscle development. A typical example of such independent development and growth is the alar and transverse atlantis ligaments at the craniovertebral junction [18]. However, the ANL and SSL appeared to be different. The present observations demonstrated that, in human fetuses, 1) proximal parts of the supinator muscle were embedded in the collagenous tissue when the developing ANL joined the thick intermuscular connecting band and 2) the anterior parts of the coccygeus muscle were surrounded by this collagenous tissue when the intramuscular tendon became the SSL. Moreover, in the ANL and SSL, muscle fibers embedded in the ligaments suggested degeneration, or at least impairment of function. In fact, macrophage accumulation has been reported in the fetal ANL at a similar stage [19]. Thus, at these two sites, ligament growth appears not to be "favorable" for the developing muscles, in contrast to the coordinated development and growth that has been hypothesized (see Introduction). The fetal supinator was very thick relative to the size of the elbow. Thus, even if the ANL remained thin during development, the supinator muscle appeared to stabilize the growing head of the radius in utero. Likewise, even if the SSL was absent, development of the fetal sacroiliac joint appeared to be not impaired. Therefore, fetal growth of the ANL or SSL may occur independently from, or even beyond, functional demand at the corresponding joint. Although muscle contraction is likely to induce initial development of the ANL and SSL, the two appear to paradoxically injure parts of the "mother" muscles.
Reidenbach and Schmidt [13] stated that the ANL develops in stages in fetuses with a CRL between 105 and 270 mm, but considered that the term "growth" was preferable to "development." In the present study, we paid close attention to the early stage of development. According to Mérida-Velasco et al. [12], the first signs of the ANL are evident at 7 weeks, and the joint capsule becomes anchored to the ligament at 9 weeks. The thick intermuscular connecting band we observed at the elbow most likely corresponded to the lateral ulnar collateral ligament in adults described by O'Driscoll et al. [20], Hannouche and Bégué [21] and Imatani et al. [22]. Thus, at 15-18 weeks, the so-called lateral collateral ligament complex of the elbow [23] seemed to develop, with subsequent thickening of the ANL. In a study involving dissection from the superficial to the deep side of the elbow including the anconeus and extensor carpi ulnaris muscles, Kim et al. [24] published many elegant images of the lateral collateral ligament complex. In spite of its rather late development, the ANL in adults became much stronger than the ligament complex. Hast and Perkins [25] reported a cadaveric elbow showing rare anomalies: the accessory supinator and tensor muscles. Because these anomalous muscles were similar to the present supinator fibers embedded in the ANL, these muscles may result from failure of the muscle fibers to become involved in the ANL.
Development of the SSL chronologically followed the formation of the initial sacrotuberous ligament. The sacrotuberous ligament appeared to begin its development as a tendon of the gluteus maximus temporarily inserted to the ischial tuberosity. In adults, the gluteus maximus is, in fact, tightly attached to the tuberosity. In contrast, the SSL is much weaker than the sacrotuberous ligament, although both are considered critical for stabilizing the sacroiliac joint [26-28]. Because of the long and loose tendinous arch of the levator ani muscle in fetuses, the growth and contraction of the fetal rectum appears to not create mechanical stress on the coccygeus and its intramuscular tendon (i.e., the primitive SSL). In any event, development of both the SSL and the sacrotuberous ligament appears to not stabilize the fetal sacroiliac joint, but occurs as a result of mechanical influence from the nearby muscles. In the present study, the intramuscular tendon of the coccygeus appeared to provide a primitive form of the SSL. The union of the fetal coccygeus muscle with the sacrococcygeus posterior muscle [16] may induce formation of the intramuscular tendon.
At the beginning of this study, we had an impression that the ANL and SSL grow in combination with degeneration of parts of the corresponding muscles on the basis of our experience in dissection of adult cadavers (see Introduction). This study did not provide evidence of muscle cell death at the interface with the growing ligament, but we demonstrated that the developing ligament involved parts of the muscle fibers. Although we had no idea whether another example is present in the human body, we believe that a negative influence of developing ligaments on the muscles in contact with them is possible.
 XML Download
XML Download