

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Macrophages play a central role in the processes of inflammation, tissue remodeling and repair in adult tissues. In inflammation, the tendon-bone interface and the synovial membrane show a common immunological response as an enthesis organ [1]. Moreover, in fetuses as well as adults, lymphangiogenesis is followed by macrophage accumulation, as macrophages themselves can transdifferentiate into lymphatic endothelium (reviewed by Kerjaschki [2]). However, in fetuses, this surprising phenomenon seemingly occurs only in loose tissue in the human fetal knee [3].
In the musculoskeletal system of embryos and fetuses, cell death with macrophage accumulation has been well described not only in the interdigital area of the foot and hand [4-6] but also in developing muscles, including those of humans (rat diaphragm [7], human quadriceps femoris muscle [8], chick neck muscles [9], murine back muscles [10] and multiple human muscles [11]). However, fetal muscle cell death is not an obligate component of morphogenesis, but a phenomenon caused by neuronal cell death and/or elimination of some muscle fiber types (reviewed by McClearn et al. [9]). Consequently, the aim of the present study was to examine comprehensively the distribution of CD68-positive macrophages in developing human extremities. We also examined the hypothetical relationship between macrophages and lymphangiogenesis. Fetal lymphatic vessels were identified with immunohistochemistry of D2-40 or podoplanin (see Materials and Methods).
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). We examined 32 extremities obtained from 8 human mid-term fetuses at approximately 11-15 weeks of gestation: 2 fetuses at 10 weeks (crown-rump length [CRL], 50 and 55 mm), 2 fetuses at 12 weeks (CRL, 90 and 95 mm), and 4 fetuses at 15 weeks (CRL, 100, 115, 125, and 130 mm). With the agreement of the families concerned, these specimens were donated to the Department of Anatomy, Chonbuk National University, Korea, and their use for research approved by the university ethics committee. Without contravening the regulations of the respective universities or hospitals, authors other than those affiliated to Chonbuk University were waived of the need to obtain permission for this research project from the corresponding committee in Japan. All fetuses had been obtained by induced abortions. After abortion, each of the mothers had been personally informed by an obstetrician about the possibility of donating the fetus for research: no attempt was made to encourage donation. Because of randomization of the specimen numbering, it was not possible to trace any of the families concerned.
The donated fetuses were fixed with 10% v/v formalin solution for more than 3 months. After division into the head and neck, thorax, abdomen and pelvis, and the four extremities, all parts were decalcified by incubating them at 4℃ in 0.5 mol/l ethylenediaminetetraacetic acid solution (pH 7.5, Decalcifying Solution B, Wako, Tokyo, Japan) for 1-3 days, depending on the size of the material. The scapula and its associated muscles were included in the upper extremity segment, whereas the hip joint was included in the abdomen and pelvis segment for the other studies. Routine procedures for paraffin-embedded histology (using sections 5 µm thick) were conducted: the left or right extremities were used for transverse sections, while the other sides were used for longitudinal sections. Parts other than the extremities were used for our recent studies, most details of which have already been published. Most of the sections were stained with hematoxylin and eosin (H&E), but some were used for immunohistochemistry as well as terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end-labeling (TUNEL).
The primary antibodies used were 1) rabbit monoclonal anti-human CD68 (1 : 100, Dako, Glostrup, Denmark) and 2) mouse monoclonal anti-human D2-40 (1 : 100, Nichirei, Tokyo, Japan). The monoclonal antibody D2-40 raised against a MW 40 KD membrane sialomucin and the molecule is identical to podoplanin that expresses specifically in the lymphatic endothelium [12]. The D2-40 antibody was used after immersion in a ligand activator (Histofine SAB-PO kit, Nichirei, Tokyo, Japan) with autoclave treatment (105℃, 10 minutes). The second antibody (Dako Chem Mate Envison Kit, Dako) was labeled with horseradish peroxidase (HRP) and antigen-antibody reactions were detected using the HRP-catalyzed reaction with diaminobenzidine (with hematoxylin counterstaining). Podoplanin immunohistochemistry was conducted on 4 larger specimens (2 specimens at 12 weeks, 2 at 15 weeks). In addition, to identify cell death with DNA fragmentation in sections of the hand and wrist, we conducted TUNEL staining using a Millipore ApopTag Plus Peroxidase in situ Apoptosis Kit (Merck, Darmstadt, Germany).
Results
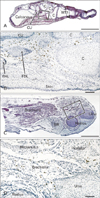
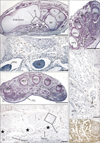
The density of CD68-positive macrophages showed a clear site-specific difference: in all specimens for all developmental stages examined, CD68-positive macrophages were concentrated in the flexor aspects of the wrist, hand and foot (Figs. 1, 2, 3, 4). In the foot, the positive cells were scattered near and along the tendons of the flexor digitorum longus and flexor hallucis longus (Fig. 1B), in the plantar intermuscular connective tissues (Fig. 2B, F), and in the sinus tarsi. The thick tendon of the peroneus longus did not appear to be associated with larger numbers of CD68-positive macrophages than those accompanying the aforementioned two flexors. In the wrist and hand, abundant positive cells were arranged along the fibrous sheath of the carpal tunnel (Figs. 2D, 3C, 4B). Some were scattered around the flexor tendons within the tunnel, as well as the narrow spaces between the carpal bone and along the interosseus muscles. However, in the extensor aspects of the extremity, CD68-positive macrophages accumulation was small and limited to the connective tissue between the carpal bones (Fig. 3B) and along the abductor pollicis brevis muscle of the wrist (Fig. 4C). Other than the sheath of the primitive carpal tunnel, no distinct ligaments carried CD68-positive macrophages in the wrist, hand and foot. The joint cavity and its lining synovium of the hand and foot did not contain CD68-positive macrophages.
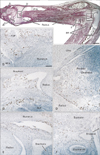
Proximal sites in the extremity contained fewer CD68-positive macrophages than the hand and foot. At the elbow, we found abundant CD68-positive macrophages in the annular ligament of the radius in 2 of 4 specimens at 15 weeks (Fig. 4E), but the medial collateral ligament did not show this feature. In the shoulder, small numbers of CD68-positive macrophages were evident along the tendons of the biceps brachii, subscapularis and/or infraspinatus muscles (figures not shown). In the knee, two of four specimens at 15 weeks carried CD68-positive macrophages in the loose connective tissue along the tendons of the hamstrings and the gastrocnemius medial head, although osteoclasts were also positive in the femur and tibia at this stage (Fig. 5B-D). CD68-positive macrophages were much more numerous in the elbow than in the knee and shoulder. In the calf, CD68-positive macrophages were concentrated in the medial intermuscular connective tissue of only one specimen (10 weeks) (Fig. 5F). None of the CD68-positive macrophages were located along arteries, veins and nerves (Figs. 2D, F, 4F and 5F), but appeared to be attached to connective tissue fibers. The joint cavity and its lining in the elbow, shoulder and knee were unaccompanied by CD68-positive macrophages (Figs. 4C, E, 5D).
Intramuscular CD68-positive macrophages were not common, but were evident in specific muscles: the distal end of the brachialis muscle at the elbow in 4 specimens (2 specimens at 10 weeks, 1 at 12 weeks and 1 at 15 weeks) (Fig. 1D), the pronator quadrate muscle in 2 specimens (1 specimen at 10 weeks and 1 at 15 weeks) (Fig. 4D), the supinator muscle (1 specimen at 10 weeks, 3 of 4 specimens at 15 weeks) (Fig. 4F), and the deltoideus muscle (1 specimen at 15 weeks) (figure not shown). Among these muscles, the brachialis muscle showed the highest number and density of CD68-positive macrophages. Intramuscular CD68-positive macrophages tended not to be concentrated either along the nerves, vessels or myotendinous junctions, but were distributed near the muscle margins or surfaces. Pathological hemorrhage, possibly resulting from the abortion procedure, was often evident in the muscles and subcutaneous layer (e.g., the subcutaneous layer shown in Figs. 2E and 5A), but few CD68-positive cells were present in the areas of bleeding. We did not examine the hip joint or its surrounding structures.
Although TUNEL staining was performed using sections of the hand and wrist of all specimens, the positive reaction was limited to tendons of the flexor digitorum profundus muscle and their associated lumbricalis muscle in the carpal tunnel (Fig. 2G). Podoplanin immunoreactivity was seen in 2 of 4 specimens examined. Most of the positive lymphatic vessels were located in the subcutaneous loose connective tissue, and did not overlap with the areas of CD68-positive macrophage accumulation (Fig. 3A). Most of the cartilages and some of the connective fibers, such as the thick sheath of the carpal tunnel, were also positive for podoplanin.
Discussion
The present study revealed a site-dependent difference in the density of CD68-positive macrophage distribution in human fetal extremities. The stage examined was much later than the stage at which interdigital apoptosis occurs in the hand and foot [4-6]. In fetuses, accumulation of CD68-positive macrophages in specific muscles has been reported [7-10] (for more details, see Introduction). In the present study also, intramuscular CD68-positive macrophages were evident in limited sites such as the distal end of the brachialis muscle. Masson trichrome staining demonstrates abundant, dark red-colored, thick muscle fibers, especially in the brachialis muscle, at 12-15 weeks of gestation (unpublished data): these muscle fibers may be in a state of degeneration. At this stage, the nerve supply to muscles has already been established. Thus, the process of muscle degeneration does not seem to target specific muscle fiber types according to nerve supply, but rather appears to represent "adjustment" or "correction" of the muscle mass (i.e., volume and shape) according to the topographical anatomy, e.g., a decrease of muscle thickness to form the terminal part of the brachialis muscle in the narrow cubital fossa. If so, this manner of development, i.e., "build and scrap," is similar to the well known mechanism responsible for forming the interdigital space at the early stage. Using human fetuses, Abe et al. [13] demonstrated that maturation of intermediate filaments in muscle fibers starts at the myotendinous junction and expands to the mid-portion of a muscle. Although the stages of muscle maturation examined in that study partly overlapped that of the present materials (10 weeks), the intramuscular CD68-positive macrophage distribution did not suggest any relationship between intermediate filament maturation and macrophages.
However, rather than in the muscle itself, the CD68-positive macrophage accumulation was more evident in loose connective tissues, especially those in the flexor aspects of the wrist, hand and foot. In these areas at these stages, multiple tendons increase in thickness, the sheath or bursa begins to develop, and re-organization of the topographical relationship between tendons may occur. Joint cavitation has already been completed. In contrast, new connective tissue spaces, such as the carpal tunnel and the sinus tarsi, appear. The new spaces in the flexor aspects are much larger than those in the extensor aspect. Macrophages seem to play a critical role in the re-organization of soft tissues at flexor sites, although details of the mechanism involved remain unknown. Sbernardori et al. [14] reported a role of mechanical stress in histogenesis of the flexor tendon pulley system in the human embryonic hand. However, they did not discuss the possible role of cell death or macrophages. The suggested mechanism of adaptation of connective tissue seems to be quite different from that observed in adult tendons or experimental models [15, 16]. Any difference in CD68-positive macrophage number between distal and proximal sites in the extremity may depend on the stage at which examination is performed: the shoulder may have higher numbers in the earlier stage than was the case in the present materials. Likewise, CD68-positive macrophages seem to play a role in maturation of the tendon-bone interface or enthesis, possibly at stages later than 15 weeks.
The present TUNEL staining of the wrist and hand revealed that positivity was limited to the flexor digitorum profundus tendon and the associated lumbricalis muscles in the carpal tunnel. The suggested cell death in the lubricalis muscle seemed to correspond to the rearrangement of deep flexor tendons for each of the fingers [17]. Thus, if cell death occurs at the other sites in and around the carpal tunnel at 10-15 weeks, it may not be due to apoptosis but rather some other mechanism, such as that postulated by Castro-Obregón et al. [18]. Developmental cell death is mediated by multiple mechanisms and displays various morphologies [19, 20]: classical apoptosis involves DNA fragmentation, whereas other forms of programmed cell death do not.
Podoplanin immunoreactivity was seen in 2 of 4 specimens examined. Most of the positive lymphatic vessels were located in subcutaneous loose connective tissue, as described previously by our group [21]. Immunoreactivity for podoplanin was also demonstrated in connective tissues by Jin et al. [22]. However, the positive fibrous tissue and lymphatic vessels did not overlap with the area of CD68-positive macrophage accumulation. Although Melrose and Little [3] reported a crucial role of macrophages in lymphangiogenesis in the human fetus at 12 and 14 weeks of gestation (completely overlapping the developmental stage of the present materials), we found no or few CD68-positive macrophages in the areas where abundant lymphatic vessels were developing. Thus it seems unli kely that, in the human fetal extremities, macrophages transdifferentiate into endothelium of lymphatic vessels. Abe et al. [23] demonstrated CD34-positive fibrous tissue in human fetal extremities, but they were located on both the extensor and flexor sides of the joints. In addition, matrix substances (aggrecan, versican, hyaluronan, and tenascin-C) do not show a specific expression pattern that overlaps with areas of CD68-positive macrophage accumulation in the hand and foot (unpublished data), although Kjaer et al. [24] and Okita et al. [25] have demonstrated changes in the extracellular matrix around tendons depending on the degree of exercise.
Finally, it seems pertinent to address individual differences in the density or immunoreactivity of CD68-positive macrophages, even though our discussion is based on general observations. Although the clinical relevance of this study is still unclear, intrauterine ultrasound may be able to evaluate changes in muscle shape and volume at 15 weeks of gestation. Morphological differences in CD68-positive macrophages are likely related to specific conditions in utero, such as general inflammation, as well as the development of muscles and tendons.


 XML Download
XML Download