

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Wnt signaling plays essential roles in morphogenesis and cellular differentiation of many tissues during development [1]. Wnt proteins transduce multiple signaling cascades including the Wnt/β-catenin pathway, the Wnt/Ca2+ pathway, and the Wnt/polarity pathway [2]. Among those, the Wnt/β-catenin pathway has been intensely studied. Wnt proteins bind to the frizzled (Frz) transmembrane receptors and the low-density lipoprotein receptor related protein (LRP5/6) co-receptors located on the cell surface to form a receptor trimeric complex, resulting in the inhibition of glycogen synthase kinase-3β (GSK-3β). β-catenin, one of the targets of GSK-3β, acts as a transcriptional co-activator of the T-cell factor (Tcf) and lymphoid enhancer factor (Lef) protein. In the absence of Wnt signaling, β-catenin is a part of cytosolic protein complex that consists of axin, adenomatous polyposis coli, and GSK-3β. GSK phosphorylates β-catenin and leads to its ubiquitination and degradation. However, the binding of Wnt protein to its receptors results in inhibition of GSK and cytosolic accumulation of β-catenin, which is translocated to the nucleus where it interacts with members of the Tcf/Lef family of transcription factors, resulting in complexes that bind to specific response sequences on the promoters of Wnt downstream target genes.
Genetic studies have shown that Wnt/β-catenin signaling plays a critical role in skeletal development and homeostasis [3, 4]. Loss of function mutations of the LRP5 leads to low bone mass accompanied by fractures causing osteoporosis pseudoglioma syndrome in humans and in animal models [5, 6]. LRP5 gain-of-function mutations in humans result in a high bone mass syndrome [7, 8]. In the gene targeted animal models, deletion of β-catenin from differentiated osteoblasts leads to osteopenia, while its stabilization in differentiated osteoblasts leads to high bone mass and failure of tooth eruption [3]. Dento-alveolar complex shares many similarities with bone. However, role of Wnt/β-catenin signaling during its formation has not been reported.
During tooth development, Wnt/β-catenin signaling plays multiple roles in various stages of tooth morphogenesis [9]. Numerous reports have shown that Wnt/β-catenin signaling is required both in the epithelium and mesenchyme for the mediation of epithelial-mesenchymal interaction during tooth morphogenesis. Inactivation of β-catenin in the dental epithelium or mesenchyme leads to arrest of tooth germs at bud stage [10, 11]. In contrast, activation of β-catenin in the dental epithelium causes multiple tooth generation [10, 12]. Activation of β-catenin in the dental mesenchyme leads to premature differentiation of dental pulp cells and produces dentin-like matrix throughout the dental pulp compartment [11]. On the basis of these findings, it is thought that Wnt/β-catenin signaling may also participate in cellular differentiation and matrix formation during tooth formation. Furthermore, Kim et al. [13] recently has found that constitutive stabilization of β-catenin in the dental mesenchyme leads to excessive dentin and cementum formation. The results suggest that local modulation of Wnt/β-catenin signaling plays critical roles in the cellular differentiation and matrix production during dento-alveolar complex formation. However, it remains unclear whether Wnt/β-catenin signaling is involved in the dento-alveolar complex formation. To understand the roles of Wnt/β-catenin signaling in the dento-alveolar complex formation, we generated and analyzed the conditional β-catenin activation mice under the control of Col1a1 promoter. Our data suggest that appropriate inhibition of Wnt/β-catenin signaling is important for the formation of dento-alveolar complex during tooth development.
Materials and Methods
Mouse strains and genotyping
All experimental procedures were approved by the Animal Welfare Committee of the Chonbuk National University. For stabilization of β-catenin in the dento-alveolar complex, Catnb+/lox(ex3) mice [14] were crossed to Col1a1-cre [15] mice. Genotypes of the mice were determined as previously described [14, 15].
Tissue preparation and histology
For histological analysis, jaws were dissected and fixed in 4% paraformaldehyde at 4℃ overnight. After rinsing with 0.01 M phosphate buffered saline (PBS), the specimens were decalcified in 10% EDTA/PBS for 2-4 weeks, then dehydrated, embedded in paraffin, and sectioned at a thickness of 7 µm. Slides were stained with hematoxylin and eosin (H&E).
Immunohistochemistry
Immunohistochemistry was performed using Histostain Plus Rabbit Primary (DAB) kit (Zymed Laboratories, San Francisco, CA, USA) and goat ImmunoCruz staining system (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) following each manufacturer's instructions. The sagittal sections were treated with 3% hydrogen peroxide, and incubated with rabbit polyclonal antibodies against β-catenin (1 : 200, Thermo Scientific, Fremont, CA, USA), dentin sialophosphoprotein (Dspp; 1 : 400, kindly provided by Dr. Larry Fisher), biglycan (Bgn; 1 : 800, Dr. Larry Fisher), dentin matrix protein-1 (Dmp1; 1 : 500, Takara Bio Inc., Shiga, Japan), phosphate regulating endopeptidase homologue on the X chromosome (Phex; 1 : 50, Sigma-Aldrich, St. Louis, MO, USA), tissue-nonspecific alkaline phosphatase (Tnap; 1 : 50, Protein Tech Group, Inc., Chicago, IL, USA), bone sialoprotein (Bsp; 1 : 1,200, Abcam, Cambridge, MA, USA), osteopontin (Opn; 1 : 500, Abcam) and goat polyclonal fibroblast growth factor-23 (Fgf23; 1 : 200, Santa Cruz Biotechnology) antibody.
Soft X-ray and micro-computerized tomographic analysis
Dissected mandibles from 4-week-old Col1a1-cre:Catnb+/lox(ex3) (MT) and wild type (WT) mice were examined on a soft X-ray system (SOFTEX CSM-2, Softex Co. Ltd., Kanagawa, Japan). After then, the mandibles were scanned in a desktop scanner (1076 Skyscan Micro-CT, Skyscan, Kontich, Belgium) and analyzed with CTscan software (Skyscan).
Body weights measurement and statistical analysis
Body weights of mice were measured at 3- and 4 weeks-old age. All statistical analyses were done using GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA, USA). Statistical differences were determined by Student's t-test and null hypotheses of no difference were rejected if P-values were less than 0.05.
Results
Tissue-specific activation of β-catenin in the dento-alveolar complex
In the mandible of mouse at P8, β-catenin was localized in the ameloblasts, odontoblasts, and osteoblasts (Fig. 1A). β-galactosidase activities were also observed in the odontoblasts and osteoblasts of the Col1a1-cre:R26R double transgenic mouse at P8 (Fig. 1B). This confirmed that targeting of β-catenin with Col1a1-cre was effective in the formation of dento-alveolar complex. At 4 week-old, height of MT mice was shorter than that of WT mice (Fig. 1C). Body weight was also decreased in the MT mice compared to the WT littermates (Fig. 1D). The mean body weight was 7.77±0.61 g (n=9) and 3.58±1.44 g (n=9) in the WT and MT mice at 3 week-old, respectively. The decrease of body weight was more evident in the 4 week-old MT mice (Fig. 1D). The mean body weight was 13.55±1.42 g (n=9) and 3.73±0.72 g (n=9) in the WT and MT mice at 4 weeks-old, respectively. The decrease of body weight was statistically significant (P<0.05). In genotyping with tail DNA, bands of 720 bps and 600 bps were amplified for cre sequences and deleted allele for β-catenin exon 3 loci (Fig. 1E). In stereoscopic observation of incisors, both of upper and lower incisors were normally erupted in the WT mice. In the MT mice, upper incisors were erupted but lower incisors were not erupted (Fig. 1F-I). Mandibular molars were normally erupted into the oral cavity in WT mice but those of MT mice were not erupted as the same as lower incisors (Fig. 1J, K).
Disturbance of tooth formation and eruption failure in Col1a1-cre:Catnb+/lox(ex3) mice
In microradiographic examination, general dimensions of craniofacial skeleton in MT mice were smaller than those in WT littermates (Fig. 2A, B). Mandible of MT mice was severely retarded in growth and mal-occluded. In microtomographic view of mid-sagittal sections of mandible, incisors were shorter and smaller than those of WT mice (Fig. 2C, D). In the molars of MT mice, root formation was severely impaired while those of WT mice were normally formed (Fig. 2E, F). In H&E stained sections, deposition of bone matrix was remarkably increased in the mandible, but was poorly mineralized in the MT mice (Fig. 2G, H). In the MT mice, molars with short roots were formed but were not erupted into the oral cavity. Overall dentin was poorly mineralized. In the consecutive sections of MT mice, β-catenin expression was increased in the odontoblasts and osteoblasts (Fig. 2I, J).
Defects of the dentin mineralization in Col1a1-cre:Catnb+/lox(ex3) mice
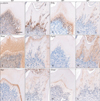
Immunohistochemistry revealed that β-catenin was localized in the differentiated odontoblasts and pulp cells of the mandibular first molar of WT mice, which is restrictedly increased in the odontoblasts of MT mice (Fig. 3A, B). Phex was localized in the differentiated odontoblasts of WT mice, but was clearly absent in the odontoblasts of MT mice except some cells included in dentin matrix (Fig. 3C, D). Dspp was extensively localized in the dentin matrix and odontoblasts of WT mice. But it was obviously decreased in the odontoblasts and predentin except thin mineralized dentin layer of MT mice (Fig. 3E, F). A mineralization inhibitor, Fgf23 was not localized in the odontoblasts as well as dentin matrix of WT mice. Interestingly, Fgf23 was extensively localized in the predentin layer of MT mice (Fig. 3G, H). Bgn was exclusively localized in the thin predentin layer in WT mice, but was significantly decreased in the MT mice (Fig. 3I, J). Dmp1 was localized in the junction area between dentin and predentin both of WT and MT mice. These results suggest that activation of β-catenin in the odontoblasts may disturb the differentiation of odontoblasts and mineralization of dentin matrix.
Root deformities and increased alveolar bone mass of Col1a1-cre:Catnb+/lox(ex3) mice
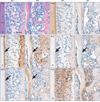
As described previously, molar roots were short and periodontium was defectively formed in the MT mice. In contrast to WT mice, root dentin was thin and hypomineralized in the MT mice. In addition, periodontal space was narrower due to excessively formed alveolar bone in the MT mice (Fig. 4A, B). In the root dentin of MT mice, Dspp was decreased in the dentin (Fig. 4C, D). In the WT mice, Bsp, Dmp1 and Fgf23 are specifically localized in the matrix of acellular cementum and alveolar bone, which was decreased in the MT mice (Fig. 4E-J). Opn was localized in the cementum and periodontal ligaments of WT mice, but it was also decreased in the cementum and periodontal ligaments of MT mice. From these results, it is suggested that activation of β-catenin in the osteoblasts and cells of periodontium may cause to increase bone formation and root deformities.
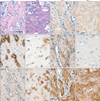
In the mandible of MT mice, trabecular spaces were reduced following increase of bone formation (Fig. 5A, B). Bsp immunoreactivity was increased whereas Tnap was decreased in the MT mice (Fig. 5C-F). Phex was strongly expressed in the bone matrix of WT mice but it was nearly absent in the MT mice (Fig. 5G, H). In the mandible of WT mice, Bgn was weakly expressed around osteoblasts. Increased Bgn immunoreactivities were observed around the osteoblasts and osteocytes in the MT mice (Fig. 5I, J). In addition, Dmp1 immunoreactivity was increased in the MT mice (Fig. 5K, L).
Discussion
It is well known that Wnt/β-catenin signaling plays critical roles in the skeletal development. Although having biochemical similarities with bone, the roles of Wnt/β-catenin signaling in dento-alveolar complex formation are unclear. Here we provide in vivo evidence that Col1a1-cre mediated activation of β-catenin leads to disturbance in tooth eruption and aberrant dento-alveolar complex formation.
Wnt/β-catenin signaling plays important roles in bone formation and maintenance. Gain- or loss-of function mutation of β-catenin in osteoblasts resulted in osteopetrosis and osteoporosis, respectively [3]. The complementary regulation of bone formation is principally controlled by the function of osteoclasts. In present data, Col1a1-cre:Catnb+/lox(ex3) mice showed disturbances in tooth eruption together with increase of alveolar bone mass. These findings indicate that persistent activation of β-catenin in the osteoblasts may cause dysfunction in osteoclasts and result in disturbance of tooth eruption. In fact, several animal models having dysfunction of osteoclasts showed tooth eruption failure together with osteopetrotic bone formation [16-20]. Therefore, it is suggested that osteoclast mediated bone resorption may be essential for potential eruption pathway of developing tooth.
In contrast to well-known function in bone formation, it remains unknown whether Wnt/β-catenin signaling is involved in the dento-alveolar complex formation. Dentin, a major component of tooth, is formed by odontoblasts differentiated from dental mesenchymal cells. It has been well known that dentin is similar with bone in its biochemical composition. However, little is known of the roles of Wnt/β-catenin signaling in dentin formation. In our data, Col1a1-cre:Catnb+/lox(ex3) mice showed aberrant dentin formation in tooth. In mutant molars, odontoblasts were not fully differentiated but prematurely differentiated following continuous activation of β-catenin. This finding is consistent with previous report that Osr2Ires-cre mediated activation of β-catenin in the dental mesenchyme leads to premature differentiation of dental pulp cells [11]. In mutant mice, Phex positive odontoblasts were scarcely found and Dspp was significantly down-regulated in the dentin matrix. In addition, Bgn, a proteoglycan in unmineralized dentin matrix, and an Fgf23, an inhibitor of mineralization, was dramatically upregulated in the predentin. These findings indicate that prematurely differentiated odontoblasts secrete organic dentin matrix but with severely impaired mineralization. From these results, it is suggested that activation of β-catenin in the odontoblasts leads to disturb the differentiation of odontoblasts and mineralization of dentin matrix. Therefore, Wnt/β-catenin signaling may positively regulate matrix production but negatively regulate mineralization during dentinogenesis.
We also demonstrate that Col1a1-cre:Catnb+/lox(ex3) mice exhibit abnormalities in dento-alveolar complex including root deformities, cementum hypoplasia and narrowing of periodontal spaces. Numerous growth factors and transcription factors such as Shh, BMP and Msx, are extensively expressed in the dental epithelium and mesenchyme during root formation [21, 22]. However, little is known about the molecular mechanisms underlying root formation. To date, several studies reported that molecular regulation is necessary for the root elongation in the dental mesenchyme. Nfic, a CTF binding transcription factor, was well known to regulate odontoblast differentiation during root formation. The Nfic knockout mice showed short molar roots [23]. In addition, it has been reported that Smad4 and Ptc1 is also associated with the root elongation [22, 24]. Ablation of these genes leads to short roots in molars together with bone defects. This implies that a common molecular mechanism may be present in the regulation of bone and tooth root formation. There are other possibilities in the disturbances of root formation in Col1a1-cre:Catnb+/lox(ex3) mice. Increased bone mass may disturb the root elongation of molars into alveolar bone during root formation. It is interesting that several animal models related to dysfunction of osteoclasts also showed short roots in molars [16-20].
Cementum also shares many similarities with bone in its biochemical compositions. The accumulating in vitro reports suggest that activation of Wnt signaling is required for the regeneration of periodontal tissue [25]. Recently, it has been shown that constitutive stabilization of β-catenin in the dental mesenchyme leads to excessive cementum formation [13]. Activation of Wnt/β-catenin signaling may stimulate cementoblast differentiation and cementum formation. In contrast, cementum formation was severely disturbed in the Col1a1-cre:Catnb+/lox(ex3) mice. This phenotypic discrepancy between two different β-catenin activation mutant lines may be related with root formation. In the Col1a1-cre:Catnb+/lox(ex3) mice, molar root formation was severely disturbed. Root defects may affect differentiation of cementoblast and cementum formation.
Our results demonstrate that local activation of β-catenin in the dental mesenchyme leads to aberrant dento-alveolar complex formation in vivo. Thus our results indicate that temporospatial regulation of Wnt/β-catenin signaling plays critical roles in cell differentiation, matrix formation and mineralization during dento-alveolar complex formation.


 XML Download
XML Download