

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
In adults, the cartilage of the pharyngotympanic tube (PTT) or simply, the tubal cartilage, covers the superior and posterior aspects of the PTT: it resembles a cap, protecting the tube against mechanical stress. However, in contrast to the tracheal and laryngeal cartilages, the tubal cartilage may bend too acutely to maintain a wide luminal space. Thus, our first motivation of this study was to consider why the tubal cartilage is not specified for the "aim." In our recent studies of fetal anatomy, including the PTT [1, 2], we have found that development of the tubal cartilage starts much later than that of the other head and neck cartilages. Although it is perhaps not widely known, the tubal cartilage begins to develop at 14-16 weeks of gestation [3], whereas the PTT develops very early. Therefore, we hypothesized that, in contrast to most of the head and neck, whose development is determined by gene cascades for regulation of the pharyngeal arch [4-7], development of the tubal cartilage is strongly dependent on later developmental events occurring along or around the PTT. Therefore, in the present study using human fetuses, we considered the converse situation of whether any problem would arise along or around the fetal PTT, if the tubal cartilage was absent.
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). We examined semiserial histological sections of 20 human fetuses at 14-18 weeks (crown-rump length [CRL], 100-150 mm) and 5 fetuses at around 30 weeks (CRL, 250-270 mm). Among the former specimens, the heads had been processed for horizontal (10 fetuses), frontal (5 fetuses) or sagittal (5 fetuses) sections. The latter 5 specimens (all sagittal sections) were used to confirm the final morphology at and around the PTT, because the morphology of the temporal bone at 14-18 weeks is quite different from that at the final stage [8]. All of the specimens were part of a large collection kept at Embryology Institute of the Universidad Complutense, Madrid, being the products of urgent abortion, miscarriages or ectopic pregnancies managed at the Department of Obstetrics of the University. Approval for the study was granted by the ethics committee of the university.
The donated fetuses were fixed with 10% v/v formalin solution for more than 3 months. After division into the head and neck, thorax, abdomen and pelvis, and four extremities, the head specimens were decalcified by incubating them at room temperature using Plank-Rychlo solution (AlCl2/6H2O, 7.0 w/v%; HCl, 3.6; HCOOH, 4.6) for 3-5 days [9]. After routine procedures for paraffin-embedded histology, sections 5 or 10 micron meter thick were prepared at intervals of 0.1-1 mm, depending on the size of the specimen. The sections included not only the PTT, but also any surrounding structures, such as parts of the brain, nose, ear and eye. All sections were stained with hematoxylin and eosin or azan.
Results
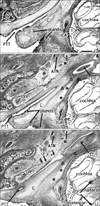
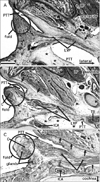
General orientation of figures
Figs. 1, 2, 3 show horizontal sections, Fig. 4 frontal sections and Fig. 5 sagittal sections. The initial appearance of the tubal cartilage is evident in Fig. 1, and a fascial connection between the cartilage and the tensor tympani muscle is evident in Figs. 1 and 2. Likewise, the topographical relationship between the cartilage and the levator veli palatini muscle is shown in Figs. 1, 3, and 4. Finally, Fig. 5 displays the pharyngeal recess. In addition, all the figures show the underdeveloped carotid canal of the temporal bone, in contrast to the late stage (Fig. 5D inset).
Initial cartilage along the PTT
In 1 of 20 specimens at 14-18 weeks, the tubal cartilage was almost round and 1 mm in diameter, being restricted to the posterior side of the pharyngeal opening of the PTT (102 mm CRL, at 14 weeks) (Fig. 1). We examined another 4 fetuses that were smaller than 110 mm (i.e., at approximately 14 weeks): The three of four specimens carried a short bar-like cartilage, extending laterally from the pharyngeal opening of the PTT (e.g., Fig. 5A, B), but the remaining specimen carried no cartilage along the PTT (Fig. 3A). Thus, it was considered that initially, the cartilage probably appeared as the medial end of the final long bar at or around 14 weeks, or at the CRL 100-110 mm stage, and then extended laterally. The initial cartilage was located at the medial end of a band-like mesenchymal condensation, which extended laterally to the tensor tympani muscle (see below).
Tensor tympani and levator veli palatini muscles
The initial round cartilage developed into a bar-like cartilage extending laterally along a fascia-like structure (Fig. 2), which first appeared as a thick band-like mesenchymal condensation (Fig. 1B, C, F, G); this early morphology was similar to a type of PTT "adventitia." However, until 18 weeks, it was visible as a fascia joining the covering fascia of the tensor tympani muscle. The cartilage-tensor tympani fascia was located on the posterosuperior side of the PTT, and the composite fibers appeared to run along the mediolateral axis. In contrast, the anterior part of the PTT adventitia connected between the tensor veli palatini muscle and Meckel's cartilage (Fig. 1C, D). Notably, during the stages examined (14-18 weeks), the tensor tympani muscle had no bony attachment because, along and near the PTT, the putative temporal bone was not developed and was restricted to the area around the cochlea (which is a part derived from the otic capsule). Thus, the medial end of the tensor tympani muscle faced, or was attached to, the lateral end of the developing PTT (Fig. 2).
The site of the initial cartilage corresponded to the immediately superior side of the developing levator veli palatini muscle (Fig. 1D, H). Before the PTT cartilage appeared, the levator muscle entered a narrow mucosal fold protruding sharply at the posterior angle of the pharyngeal opening of the PTT (Fig. 3A). Later, depending on the development of the cartilage, the mucosal fold became thick with glands (Fig. 3B, C). Notably, the levator muscle used the mucosal fold for downward passage to the palate: this was another reason for the fold becoming thick. However, the salpingopalatine fold was not evident in the fetuses examined. Notably, instead of the bony attachment, the superior end (the putative origin from the temporal bone) of the levator muscle originated from a thick fascia, whose fiber components ran in the mediolateral direction along the inferior aspect of the PTT (Fig. 4). This inferior fascia was separated from the other peritubal fascia, which was connected with the tensor tympani muscle, but in both, the composite fibers ran along the mediolateral axis (see the paragraph above). Moreover, part of the levator muscle was attached to the cartilage itself (Fig. 4B, C). On the medial side, the levator muscle issued a fascia that reached the anterior aspect of the longus capitis muscle (Fig. 3C). In addition, the tensor veli palatini muscle issued a fascia that extended laterally to Meckel's cartilage (Figs. 1C, 2C), as well as medially to the pterygoid process of the sphenoid bone (Fig. 3A, B). However, the fascia of the tensor veli muscle did not connect with the tubal cartilage at the early stage. Likewise, the origin of the tensor veli muscle from the lateral lamina of the tubal cartilage was not evident in the early stage of development. The tensor veli palatini muscle originated from the pterygoid process, which had already started to ossify at 14 weeks.
Carotid canal and other structures
At 14-18 weeks, the internal carotid artery ran superiorly along the anterior surface of the cartilaginous cochlea. Thus, the artery was exposed to loose mesenchymal tissue, facing the PTT and tubal cartilage (Figs. 1, 2, 3, 5B, C); this tissue mass, or a loose space, was located on the posterosuperior side of the PTT (Figs. 2-5). Notably, in the large fetuses examined, the space was completely occupied by the later-developing part of the temporal bone; thus, the bony carotid canal was established (Fig. 5D inset), and the loose space was replaced by the anterior wall of the bony carotid canal. Subsequently, the levator veli palatini muscle acquired its bony attachment to the inferior extension of the temporal bone.
The pharyngeal recess, or Rosenműller's fossa, of the pharyngeal cavity was clearly identifiable in sagittal sections (Fig. 5A, C). The PTT cartilage was sandwiched between the recess and the PTT. In horizontal sections, the recess corresponded to part of the pharyngeal cavity behind a large mucosal fold containing the cartilage and levator muscle (Fig. 3C). The shape and size of the pharyngeal recess did not show individual variation in the fetuses examined, in contrast to the observations of adults by Khoo et al. [10]. The salpingopharyngeal fold was not evident in fetuses, but the salpingopharyngeal muscle was identified as a medial part of the constrictor pharyngis superior muscle (Fig. 4).
Discussion
The present study demonstrated that the tubal cartilage developed along a peritubal fascia connecting with the tensor tympani muscle. Likewise, the levator veli palatini muscle originated from another peritubal fascia on the inferior side of the PTT. Because their composite fibers were oriented in the same mediolateral direction, these peritubal fasciae were likely to be mutually continuous in the posterior aspect. In addition, a similar peritubal fascia was evident between the tensor veli palatini muscle and Meckel's cartilage. Thus, instead of the later-developing bony attachments, the PTT provided fascial structures for all of the three paratubal muscles. The peritubal fasciae seemed to develop as a migration fascia against a growing structure (i.e., the PTT), being apparently analogous to the renal fascia against the developing kidney [11, 12]. However, unlike the renal fascia, the paratubal muscles used the fasciae as an origin at the early stage. The fascia for the levator veli palatini muscle origin extended medially to the prevertebral fascia covering the longus capitis muscle. Miyake et al. [13] reported that the latter develops as a kind of intermediate tendon between the bilateral prevertebral muscles. Thus, multiple fasciae with different origins might be interconnected.
In adults, it is well known that the tensor veli palatini, as well as the tensor tympani, becomes attached to the tubal cartilage [14]. However, the tensor veli muscle origin from the lateral lamina of the tubal cartilage was not evident during early development. It has been noted that the levator veli in both the adult and the newborn originates via a cylindrical tendon attached to the inferior surface of the petrous part of the temporal bone near the carotid foramen, and also to the tubal cartilage [15]. However, instead of the petrosal part, a loose space was noted between the internal carotid artery and the PTT at 14-18 weeks. Klueber and Langdon [16] reported that, in 15-week human fetuses, the levator veli muscle sometimes originates from the membranous tube, although they did not describe the peritubal fasciae. We observed the tubal cartilage as a limited hard tissue that was available for the origins of the developing levator veli and tensor tympani muscles. Conversely, via the peritubal fasciae, the tubal cartilage at the early stage seemed to receive mechanical stress for growth. The tensor tympani muscle appeared likely to lead or induce lateral extension of the cartilage along the fasciae, whereas the levator veli muscle was likely to provide cues for extension of the medial lamina of the cartilage. Therefore, the tubal cartilage seems to start development after the establishment of paratubal muscles. The PTT developed first, paratubal muscles second, and the tubal cartilage third.
Swarts et al. [3] reported that the distances of the levator veli and tensor veli muscles from the tubal lumen remain essentially unchanged throughout development, when standardized relative to the changes in body size. Moreover, the topographical anatomy of the PTT and paratubal muscle origins in normal newborns and those with cleft palate is almost the same [17]. Such stable morphology of the paratubal muscles seemed to be a result of the early development, in accordance with the PTT. Subsequently, the tubal cartilage morphology is also stable, although after birth, the shape begins to show a difference between the normal situation and that of the cleft palate [18]. Mallo et al. [19] demonstrated a critical role of the epithelium in the external ear cartilage pattern formation. In fact, the initial cartilage bars develop along the external auditory meatus, and surround it (now submitted to Ann Anat). However, development of the tubal cartilage might not depend on the differentiation of the PTT epithelium, including the glands. Rather than having a positive influence, the glands often invaded into the tubal cartilage, and sometimes divided the latter, as has been reported in the epiglottic cartilage [2]. The invasion of glands may be related to the distribution of elastic fibers, those developed in the late stages in the fetal tubal cartilage (unpublished data).
On the basis of our findings and the available data, we hypothesize that 1) one, the PTT and paratubal muscles differentiate through predetermined gene cascades that are responsible for development of the first and second pharyngeal arches; 2) the other, the tubal cartilage mechanically supports the development of paratubal muscle, and vice versa.

 XML Download
XML Download