

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Animals can detect temperature changes in their environment with special thermoreceptors, which aid in hunting, feeding, and survival in many organisms, including crotaline and boid snakes [1], vampire bats [2, 3], fire-seeking beetles [4, 5], certain butterflies [6, 7], and blood-sucking bugs [5, 8]. Crotaline and boid snakes have specialized infrared (IR) receptors in pit organs that enable them to detect, locate, and apprehend prey when combined with other sensory systems [1]. IR sensitivity in crotaline and boid snakes has likely evolved from the somatic sensory system, which evolved to sense IR radiation, similar to the vision sensation mechanism [9]. IR radiation sensitivity is much greater in crotaline and boid snakes than in other thermosensitive animals, and crotaline snakes have more sensitive IR radiation receptors than boid snakes [10].
This review describes the structures of the IR-sensitive pit organ and trigeminal sensory systems, which connect IR sensation to the brain, in crotaline snakes.
Pit Organ Location and Structure in Crotaline Snakes
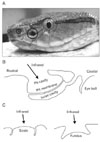
Two snake families have IR receptors: Boidae (boid snakes) and the subfamily Crotalinae in the Viperidae (crotaline snakes). Crotaline snakes have pits comprising the pit organ located on both sides of their face roughly midway between the nostril and the eye [1, 11]. The pit organ of crotaline snakes consists of a 1-5-mm-wide cavity, several times the diameter of the nostril (Fig. 1A) [11]. A thin pit membrane is suspended inside the cavity between the inner and outer chambers of a pair of pits in the loreal region [11]. The outer chamber opens widely to the exterior, whereas the inner cavity interacts with the external air via a sphincter-controlled pore rostral to the eye [11]. Fig. 1B shows a diagram of a crotaline pit organ in cross-section. In comparison, boid snakes have receptors in their labial scales, either without specialized structures or in the fundus of specialized labial pits (Fig. 1C) [1, 10, 12].
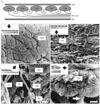
IR radiation sensitivity is much greater in crotaline snakes than in boid snakes [10, 12, 13], and IR sensation in crotaline snakes requires lower temperature changes and energy thresholds of 0.003℃ and 10.75 µW/cm2, respectively, and has a greater detection distance (66.3 cm) than that in boid snakes [10, 13], due to receptor depth and pit organ anatomy [9]. In the crotaline snake Trimeresurus flavoviridis, the pit organ membrane is approximately 15 µm thick [11] and is surrounded by air on both sides, preventing heat loss by conduction to surrounding tissues and allowing most of the IR radiation incident on the pit to heat the membrane [11]. Approximately 7,000 IR-sensitive trigeminal sensory axon endings are distributed throughout the membrane, which excite their nerve fibers when warmed [9]. Similar heat-sensitive nerve endings cover the bottom of each pit in boid snakes [9], because their receptors are not suspended, as in crotaline snakes. Unlike the receptors in the floor of the labial pits or scales, the pit membrane of crotaline pit organs is highly innervated and has a dense capillary bed that projects throughout the network of mitochondria-rich terminal nerve masses (TNMs), supplying blood for cooling and providing energy and oxygen to the TNMs (Fig. 2A) [9, 14]. The pit membrane vasculature is finer, flatter, and more convoluted than that of other sensory organs, including the skin and retina [14]. Scanning electron microscopy has revealed the three-dimensional morphology of the fundamental structures that comprise the IR receptor system in crotaline snake pit membranes (Fig. 2). The TNMs are arranged in one layer directly beneath the outer epithelium of the pit membrane (Fig. 2A, B). A group of myelinated nerve fibers branch to innervate the TNMs at a point farthest from the outer epithelium, below the TNMs and capillary network (Fig. 2A, C). The intimate apposition of the blood capillaries with the TNMs and their feeder nerve fibers is shown in Fig. 2D. Viewed from the inner chamber, the myelinated fibers are beneath the TNMs and become unmyelinated as they bend upward to form the array of sensors (Fig. 2E).
In crotaline snakes, the pit organ has unusual vasculature and numerous IR-sensitive TNMs [11, 14, 15]. Due to the extensive nature of the vasculature and its proximity to the TNMs [14, 15], chemicals administered via the blood reach the TNMs. This morphology has been examined by studying the IR receptor response to vasoactive chemicals, which occurs either by a vasoactive effect on the pit membrane vasculature or by a chemical effect on the IR receptor thermoreceptor channels, such as the transient receptor potential (TRP) vanilloid family [16-18]. Gracheva et al. [19] and Panzano et al. [20] found that pit organs respond to temperature using the heat-activated cation channel TRP ankyrin 1 (TRPA1). They suggested using TRPA1 and other TRP channels as new genetic and physiological markers to delineate evolutionary relationships between vertebrates and invertebrates. Further studies are required to confirm if IR receptors have other TRP channels.
The Trigeminal Sensory System Connects IR Sensation to the Brain in Crotaline Snakes
In mammals, the trigeminal nerve connects sensations including touch, pressure, pain, and temperature from the facial, maxillary, and mandibular regions to the brain. Crotaline snake pit organs are also innervated by the trigeminal nerve, which contains a very pure population of unimodal warm nerve fibers [1]. Three branches of the trigeminal nerve innervate the pit membrane: one ramus ophthalmicus branch and two ramus maxillaries branches. These trigeminal ganglion (TG) neuron fibers connect ipsilaterally in the lateral descending nucleus of the medulla oblongata [21, 22]. This system is independent of the common trigeminal sensory system found in these snakes and other vertebrates. Other reports have examined the IR afferent system, including the nucleus reticularis caloris [23] and optic tectum [21, 24].
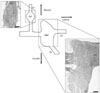
In amphibians and mammals, the ophthalmic and maxillomandibular ganglia are fused to form one complex trigeminal or semilunar ganglion. Conversely, the two ganglia remain separate in most reptiles [25]. A prominent septum further separates the two divisions of the maxillomandibular ganglion in crotaline snakes [26]. In a horseradish peroxidase (HRP) tracing study, HRP cell labeling from the pit organ to the TG revealed HRP-positive IR-sensory TG neurons in the ophthalmic ganglion and the maxillary division of the maxillomandibular ganglion but not in the mandibular division [27]. Fig. 3 shows a diagram of the TG structure in crotaline snakes. Based on its specialized structure, several immunohistochemical studies have described the distribution of various neuronal signals in the TG, including calcitonin gene-related peptide [28], neuronal and endothelial nitric oxide synthases [29, 30], and protein kinase C delta [31].
Concluding Remarks
Some animals and insects have thermoreceptors, which aid hunting, feeding, and survival. IR radiation sensitivity is much greater in crotaline and boid snakes than in other thermosensitive animals. Furthermore, crotaline snakes have a more sensitive IR receptor than boid snakes because they have a thin thermosensitive membrane suspended between a pair of pits, located in the loreal region, which comprises the pit organ. The pit membrane vasculature, located near IR-sensitive terminal nerve masses, the IR receptors, supplies blood for cooling and provides the energy and oxygen required by the IR receptors. The ophthalmic and maxillary branches of the TG nerves innervate the pit membrane. In crotaline snakes, the TG is separated into the ophthalmic and maxillomandibular ganglia and a prominent septum further separates the two divisions of the maxillomandibular ganglion. TG ganglion neurons in the ophthalmic ganglion and the maxillary division of the maxillomandibular ganglion, but not in the mandibular division, connect IR sensation to the brain. Further studies are required to expand what is known about the thermosensitive TRP channels in the crotaline pit organ IR receptor.
 XML Download
XML Download