

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Vitamin D3 (1, 25-dihydroxyvitamin D3) has been associated with the regulation of Ca2+ and Pi transport and bone mineralization [1, 2]. This association has been supported by vitamin D3-related gene knockout studies [3-5]. However, recent studies indicate that vitamin D3 affects cell proliferation, differentiation, and gene regulation [2]. Moreover, three kinds of evidence suggest an association between vitamin D3 and carcinogenesis. The first class of evidence is derived from epidemiological studies. A low serum vitamin D3 level has been associated with an increased risk for colorectal [6], breast [7], and prostate cancers [8]. Moreover, Giovannucci et al. [9] studied the relationship among dietary and supplementary vitamin D, physical activity, body mass index, and sunlight exposure in 47,800 individuals and found a strong association between low levels of vitamin D3 and increased cancer incidence, particularly for cancers of the digestive system. The second class of evidence is derived from altered expression of vitamin D3-related genes in many tumor types. Overexpression of CYP24, which downregulates the level of the active form of vitamin D3, has been reported in various cancers, including colon [10, 11] and esophageal [12] cancers. Amplification of 20q13.2, which harbors the CYP24 gene, has been reported in gastric and breast cancers [13, 14]. The third class of evidence is derived from studies about vitamin D3-related gene knockout mice. In vitamin D receptor knockout mice, a higher incidence of lymphoblastic and thymic lymphoma and mammary hyperplasia is observed [15].
Gastric cancer is the second leading cause of cancer-related deaths in the world [16]. Gastric cancer is a malignant tumor that originates from gastric mucosa and is classified into intestinal and diffuse types. Many factors are related to the cause of gastric cancer, such as a high-salt diet, ingestion of grilled meats and fish, and infection by Helicobacter pylori [17]. These bacterial, environmental, and host factors contribute to the molecular changes of gastric cancer. Despite advances in diagnostic technology such as endoscopy, the cure rate for gastric cancer is still poor due to fast growth and metastasis. Cholangiocarcinoma is a malignant tumor that originates from bile duct epithelial cells [18]. Intrahepatic cholangiocarcinoma is the second most common subtype of primary hepatobilliary cancer [18, 19]. Chronic inflammation of the liver contributes to the malignant transformation of cholangiocytes. The main reasons for the poor prognosis of patients with cholangiocarcinoma are due to diagnostic difficulty, extensive local tumor invasion, and multidrug resistance. Many molecular factors contribute to carcinogenesis of gastric cancer and cholangiocarcinoma [20-23]. Interestingly, hedgehog (Hh) signaling contributes to the progression of both cancers [24].
Although vitamin D3 regulates the growth of various cancers, vitamin D3 has not been examined in gastric cancer and cholangiocarcinoma. Herein, we report that vitamin D3 inhibits cell viability in gastric cancer and cholangiocarcinoma cells.
Materials and Methods
Cell culture
SNU1, SNU638, and SNU1079 cells were cultured with RPMI-1640, 25 mM HEPES, 10% fetal bovine serum (FBS), and 1× penicillin/streptomycin. HuCCT1 cells were cultured in RPMI-1640, 10% FBS, and 1× penicillin/streptomycin at 37℃ in a 5% CO2 incubator. SNU1, SNU638, and SNU1079 cells were purchased from the Korea Cell Line Bank (Seoul, Korea). HuCCT1 cells were purchased from the Health Science Research Resources Bank (Osaka, Japan). Vitamin D3, cyclopamine, paclitaxel, adriamycin, and vinblastine were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cell viability assay
Cells were seeded at a density of 1×103 (SNU1 and SNU638), 1.5×103 (HuCCT1), or 4×103 cells (SNU1079) per well in 96-well plates and treated with the indicated concentrations of vitamin D3. Four days following treatment, 10 µl of Ez-Cytox reagent (ITSBio, Seoul, Korea) was added to the wells, and the cells were incubated for an additional 2 hours under normal cell culture conditions. Cell viability was measured by absorbance at 450 nm using an ELISA reader (Tecan, Mannedorf, Switzerland).
Real-time polymerase chain reaction (PCR)
Total RNA was extracted using a RNeasy Mini kit (Qiagen, Valencia, CA, USA) and processed based on the manufacturer's manual. cDNA was synthesized with MMLV reverse transcriptase (Promega, Madison, WI, USA), dNTP, and oligo-dT primers. Real-time PCR was conducted using FastStart Universal SYBR Green Master Mix (Roche Applied Science, Indianapolis, IN, USA) in an ABI Prism 7500 sequence detector (Applied Biosystems, Foster City, CA, USA). The primer sequences were as follows: Ptch1 (F: 5'-AAC CCC TGG ACG GCC GGG AT-3', R: 5'-AGG ATG ACC ACG GGC ACG GCA-3'); Gli1 (F: 5'-TGC TGG ATG GGC GGG AGG ACC-3', R: 5'-CCC CGT GGA TGT GCT CGC TGT-3'); CCND1 (F: 5'-CGC GCA GAC CTT CGT TGC CCT-3', R: 5'-GCC TTG CAC TGC GGC CAC CA-3'); Bcl2 (F: 5'-CTG GGG GAG GAT TGT GGC CTT CTT TG-3', R: 5'-TCC AGG TGT GCA GGT GCC GGT TC-3'); and β-actin (F: 5'-GCG AGC ACA GAG CCT CGC CT-3', R: 5'-GCC TTG CAC ATG CCG GAG CC-3').
Data analysis
All data are presented as mean±SD. All experiments were repeated at least four times. The difference between the mean values of the two groups was evaluated using the Student's t-test (unpaired comparison). We used a one-way analysis of variance, followed by Tukey's multiple comparison to compare more than three groups. A P-value<0.05 was considered statistically significant.
Results
Vitamin D3 inhibits viability of gastric cancer and cholangiocarcinoma cells
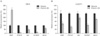
We used SNU1 and SNU638 cells to test the effect of vitamin D3 on the viability of gastric cancer cells. Cells were seeded in 96-well plates and treated with different concentrations of vitamin D3 in the presence of 1% FBS. After 4 days, we measured the absorbance. Vitamin D3 treatment significantly inhibited the viability of gastric cancer cells compared to that in control cells in a dose-dependent manner (Fig. 1A, B). Specifically, treatment with 10 µM vitamin D3 suppressed viability by >80%. Subsequently, we tested the effect in HuCCT1 and SNU1079 (cholangiocarcinoma) cells. Vitamin D3 treatment inhibited viability of cholangiocarcinoma cells as in gastric cancer cells (Fig. 1C, D).
Vitamin D3 shows a synergistic effect with other anti-cancer drugs
Brüggemann et al. [25] reported that vitamin D3 does not exert a synergistic effect with other anti-cancer drugs. However, it remains unclear whether or not vitamin D3 has a synergistic effect with other anti-cancer drugs in gastric cancer and cholangiocarcinoma cells. Thus, we seeded SNU1 and HuCCT1 cells in 96-well plates and added vitamin D3 with anti-cancer drugs, such as paclitaxel, adriamycin, and vinblastine in the presence of 10% FBS. Cell viability was measured after 2 days of treatment. As shown in Fig. 2, a significant difference was observed in cell viability between the vitamin D3 alone and the combined with anti-cancer drugs groups. Treatment with vitamin D3 alone decreased the average survival rate of SNU1 and HuCCT1 cells by 57% and 41% respectively (Fig. 2). However, combined treatment with other drugs decreased the survival rate by >70%. This result indicates that vitamin D3 can function synergistically with other anti-cancer drugs.
Vitamin D3 inhibits the expression of Hh signaling target genes
To investigate the underlying mechanism involved in the regulation of viability by vitamin D3, we examined Hh signaling because cyclopamine, a specific inhibitor of the Hh signaling pathway, almost completely suppressed cancer cell viability (Fig. 1). Furthermore, Bijlsma et al. [26] reported that vitamin D3 is an antagonist of Hh signaling. However, the relationship between vitamin D3 and Hh signaling is not well-studied in cancer cells. To determine whether or not vitamin D3 is associated with Hh signaling in cancer cells, we assessed Hh signaling activity by measuring the Ptch1 and Gli1 expression levels, which are direct target genes of Hh signaling. SNU1 and HuCCT1 cells were treated with 10 µM vitamin D3 one day after cell seeding. One day following the treatment, the Ptch1 and Gli1 mRNA expression level decreased sharply (Fig. 3). We also examined effects on the expression of other Hh signaling targets, including cyclin D1 and bcl2. In particular, we have shown previously that inhibiting Hh signaling reduces bcl2 expression in gastric cancer cells [27]. Vitamin D3 treatment reduced cyclin D1 and bcl2 expression (Fig. 3). These results suggest that vitamin D3 may act as an antagonist of Hh signaling in gastric cancer and cholangiocarcinoma cells.
Discussion
Vitamin D3 has been widely used clinically, and the proper dosage and side effects have been well-documented [28]. In the present study, we showed, for the first time, that gastric cancer and cholangiocarcinoma cells are sensitive to vitamin D3, and that vitamin D3 has synergism with other cancer drugs. Moreover, we also showed that the actions of vitamin D3 are possibly mediated through inhibition of Hh signaling.
The expression of Hh signaling target genes was regulated by vitamin D in the present study (Fig. 3). Hh signaling is one of the most important signaling pathways regulating proliferation, differentiation, embryogenesis, and cancer [29]. In mammals, Hh consists of three different members, including Sonic hedgehog, Indian hedgehog, and Desert hedgehog [30]. The trigger for signaling is the Hh ligand binding to a 12-pass transmembrane protein patched (Ptch1), which resembles the Niemann-Pick disease type C1 (NPC1) protein. The NPC1 protein is involved in cholesterol trafficking and has a pump function [26]. After Hh binds to Ptch1, Ptch1 no longer blocks the 7-pass transmembrane protein, smoothened (Smo), and subsequently the intracellular signaling cascade is activated, including Fused, protein kinase A, GSK3, CKI, and Gli. Finally, the Gli protein is activated and translocated to the nucleus where it turns on target genes. However, in the absence of Hh, Smo is continuously repressed by Ptch1. Finally, the Hh target genes are turned off.
The association between vitamin D and Hh signaling was first reported by Bijlsma et al. [26]. Bijlsma et al. [26] reported that Ptch1 can secrete 3β-hydroxysteroids, which inhibit Smo on other cells. Vitamin D3 directly binds to Smo with high affinity in a cyclopamine-sensitive manner. Moreover, Bijlsma et al. [26] showed that treating zebrafish embryos with vitamin D3 mimics the Smo-/- phenotype. Their report suggested that vitamin D3 can be used as an anti-cancer drug. In contrast, Brüggemann et al. [25] reported that vitamin D3 inhibits pancreatic cancer cell viability in the presence of 0.5% serum; however, it does not inhibit the pancreatic cancer cell viability in the presence of 10% serum. Moreover, vitamin D3 did not show any synergism with other anti-cancer drugs for inhibiting cell viability. Brüggemann et al. [25] suggested that vitamin D3 is not an efficient anti-cancer drug. However, in the present study, vitamin D3 had the ability to inhibit cell viability and showed a synergistic effect with other anti-cancer drugs in 10% serum, as shown in Fig. 2. Hence, our data indicate that vitamin D3 could be used as an anti-cancer drug, and its effect may depend on cell type.
Vitamin D3 is related to cancer via various signaling pathways [2]. However, in the present study, we focused on Hh signaling because cyclopamine, a specific inhibitor of Hh signaling, almost completely suppressed cancer cell viability, which was consistent with previous reports [24]. Although it is unclear how the Hh signaling pathway is activated in gastric cancer and cholangiocarcinoma, it is clear that Hh signaling activity is critical for viability [24, 27]. We showed that vitamin D3 reduced the expression of Hh signaling target genes (Fig. 3), suggesting the possibility that Hh signaling may play a critical role in vitamin D3-induced inhibition of viability. Future studies such as Gli1 overexpression are required to confirm this possibility.


 XML Download
XML Download