

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Clusterin (CLU) was found initially as a cell-aggregating factor in ram rete testis fluid [1]. Two decades ago, CLU, a glycoprotein also known as apolipoprotein J, testosterone-repressed prostate message-2, and sulfate glycoprotein-2, was first cloned and characterized as a 427 amino acid polypeptide, which is post-translationally cleaved between residues 205 and 206 [2, 3]. The CLU transcript is expressed relatively highly in the brain, ovary, testis, and liver, and less abundantly in the heart, spleen, lung, and breast [3]. CLU was soon suggested as a marker of cell death, as it is upregulated in many cell types following cytotoxic stimulation [4-7]. However, accumulating results have revealed that CLU is a much more complicated protein than initially thought. In this review, we explore the pro-apoptotic role of nuclear CLU (nCLU) in vitro and in vivo in the context of brain damage and neurodegeneration.
Without glycosylation, human CLU is a protein of approximately 50 kDa and detected as a ~60 kDa glycosylated precursor secretory CLU (psCLU). It is cleaved to α and β chains of ~40 kDa and further glycosylated to form mature disulfide-linked heterodimeric secretory CLU (sCLU, 75-80 kDa). The nCLU transcript lacks the endoplasmic reticulum (ER)-targeting sequences at exon 2 and utilizes the second ATG site at exon 3 (Fig. 1A). The product is detected as the ~49 kDa nonglycosylated precursor nCLU in the cytosol and as a ~55 kDa glycosylated protein (nCLU) in the nucleus [8]. The CLU protein has two coiled coil domains responsible for interacting with other proteins, e.g., Ku70, which binds to CLU upon DNA damage [9] and two nuclear localization signals [7].
Proapoptotic Role of nCLU
CLU plays controversial roles in apoptosis by producing two alternatively spliced isoforms in various cell types [10]. The pro-apoptotic CLU appears to be nCLU [7], and the sCLU and intracellular CLU are thought to be anti-apoptotic [11, 12]. The dichotomous roles of CLU in cell death were obvious from early studies using transgenic and gene-targeted mice, in which overexpression and deletion of the genes both caused reduced brain damage following hypoxia in vivo [13, 14]. Furthermore, previous reports showed that healthy human prostate cells expressed nCLU exclusively, but that human prostate cancer cells lost nCLU expression while increasingly expressing sCLU upon metastasis [12] and that deficiency of Clu enhances prostate cancer metastasis in prostate cancerprone transgenic adenocarcinoma of mouse prostate mice [15]. CLU translocation from the nucleus to the cytoplasm is related directly to colon tumorigenesis [16]. Considering these results, the expression and function of CLU in vivo must be finely tuned at the transcriptional, translational, and posttranslational levels.
As mentioned above, CLU is a bifunctional protein in terms of cell death and survival; sCLU or intracellular CLU inhibits apoptosis by interacting with Ku70 and Bax [17], whereas nCLU induces cell cycle arrest and cell death by inhibiting nuclear factor-κB-dependent Bcl-XL expression [18, 19]. In contrast to a previous study, in which intracellular CLU inhibited apoptosis by interacting with activated Bax [11], our group recently reported that nCLU sequestered Bcl-XL via a putative Bcl-2 homology 3 (BH3) domain, which was demonstrated in cells transfected with CLU gene deletion mutants [20] and by nuclear magnetic resonance spectroscopy [21]. Subsequently, Bax is released from Bcl-XL, promoting apoptosis accompanied by caspase-3 activation and cytochrome c release, implying that the role of nCLU is similar to derepressor/sensitizer BH3-only proteins [20].
BH3-only proteins are pro-apoptotic proteins with a single BH3 domain, unlike other Bcl-2 proteins that have multiple BH domains. Although the precise mechanism of the function of BH3-only proteins remains elusive, BH3-only proteins appear to interact with anti-apoptotic Bcl-2 proteins, e.g., Bcl-2 and Bcl-XL, and to activate effector Bcl-2 proteins, such as Bax and Bak, directly as an activator or indirectly as a derepressor/sensitizer, leading to cytochrome c release from mitochondria and cell death [22-27]. Overexpression of BH3-only proteins promotes apoptosis in many cell types but requires either Bax or Bak as an effector [28]. There are many BH3-only proteins, such as Bid, Bim, Bad, Bmf, Bik/Blk, Hrk, NOXA, and PUMA [29, 30]. Notably, BH3-only proteins do not usually have any other known functional domains rather than the BH3 domain [31]. In human nCLU, a putative BH3 motif was found in a C-terminal coiled coil (CC2) domain (Fig. 1A) [20, 21]. The sequence and tertiary structure of the putative BH3 motif were comparable to those of previously known BH3-only proteins (Fig. 1B). In particular, the hydrophobic L323 residue (in red Fig. 1B) and uncharged N328, which are highly conserved in BH3-only proteins, are indispensable for binding to Bcl-XL [20, 21]. Chemical shift perturbation data suggest that the nCLU BH3 domain could bind Bcl-2 as well [21], but that the actual interaction with Bcl-2 appears to be weaker than that of Bcl-XL [20].
We have also found that only nCLU, not sCLU, interacts with Bcl-XL [32]. Cleavage and heterodimerization to form sCLU could change protein folding and conformation, which might explain the differential roles of sCLU and nCLU in binding to Bax and Bcl-XL, respectively, and eventual cell survival/death. A recent report showed that the binding motif in psCLU was localized in the disulfide constraint region and was essential to bind IκB-α, but neither the α- nor the β-chain of sCLU had any activity [33]. This is another example to understand the importance of CLU protein conformation and tertiary structure. In addition, post-translational modifications could further regulate the differential roles of CLU. It is known that nCLU, located in the nucleus, is not glycosylated [34], whereas mature sCLU is heavily glycosylated [35].
CLU in Brain Damage and Neurodegeneration
CLU is expressed during fetal and postnatal central nervous system (CNS) development in mice [36]. CLU expression is upregulated in rats following traumatic brain injury [37], in seizure [38, 39], and in some neurodegenerative diseases, such as Parkinson's disease (PD) and Alzheimer's disease (AD) [40, 41]. The role of CLU in brain cell death is contradictory, as both gene-deficiency and overexpression of CLU inhibit brain damage in mice [13, 14]. It should be noted that the experimental conditions were different, as neonatal hypoxia/ischemia and permanent ischemia by middle cerebral artery occlusion in adult mice were used, respectively. CLU increases post-ischemic cell death in hippocampal slice cultures [42], implying a pro-apoptotic role for CLU.
We have discovered that nCLU plays a pro-apoptotic role in ethanol-induced cell death in the developing brain of infant rats [32]. We found that CLU increased in the cerebral cortex following acute ethanol treatment. CLU is broadly upregulated in the cerebral cortex, such as in Layers II, IV, and VI, and in the retrosplenial granular (RSG) cortex and the amygdala at 24 hour post-ethanol treatment. This CLU upregulation is related to increased cell death, which was assessed by activated caspase-3 levels and TUNEL staining. The upregulated CLU is a nuclear isoform that is translocated to the nucleus upon ethanol exposure and interacts with Bcl-XL, mediating apoptosis.
CNS dysfunction is one of the characteristics of fetal alcohol spectrum disorder (FASD), accompanied by abnormal brain structure/function, reduced white and grey matter volumes, and reduced total brain volume [43, 44]. It should be mentioned that our experimental model was heavy binge drinking, rather than chronic alcoholism. CLU expression was upregulated in the dispersed areas of the cerebral cortex, such as Layers II, IV, and VI, and in the amygdala upon acute ethanol exposure [32]. The amygdaloid complex is composed of more than ten nuclei, and one of the major functions of the amygdala in rats and humans is enhanced memory formation for emotional stimuli [45]. Upregulated CLU expression was found in the RSG cortex as well. The RSG cortex is an important link between hippocampal formation and other limbic areas of the brain and is equivalent to the cingulated gyrus in humans [46, 47]. It is also part of "Papez circuit," which was proposed by Papez as an emotion system [48]. The prefrontal cortex is involved in higher brain functions, including cognitive functions and behavioral control [49]. Furthermore, CLU upregulation is accompanied by upregulation of activated caspase 3 and TUNEL positivity in the brain following ethanol exposure, implying a role for CLU in brain cell death [32].
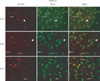
Additionally, we have found CLU upregulation and an interaction between CLU and Bcl-XL in the thalamus of the developing brain in infant rats upon ethanol exposure. As seen Fig. 2, CLU level increased post-ethanol exposure (left column), and CLU and Bcl-XL were co-localized (right column). Recent studies have reported that ethanol increases activated caspase-3 and subsequent neuronal cell death [50], which may be a result of ER stress induced by ethanol [51]. Nevertheless, the effect of ethanol on the thalamus is less understood. Our results suggest that CLU upregulation and interaction with Bcl-XL may mediate thalamic cell death by ethanol. The thalamus is located deep inside the forebrain and is often referred to as the gateway to the cerebral cortex, as thalamic neurons send axons to the cortex via the internal capsule [48]. In general, the internal capsule axons carry information to the cortex about the contralateral side of the body. It would be interesting to know the potential effect of ethanol on CLU expression and the interaction with other molecules and mechanisms thereof in the thalamus to understand the cause of behavioral deficits of FASD. Collectively, brain cell death mediated by nCLU in those areas might result in behavioral deficits and CNS abnormalities manifested in FASD. Furthermore, it is possible that CLU may be upregulated in other brain areas or cell types as well, such as the cerebellum, hippocampus, or microglial cells, which are known ethanol targets [52, 53]. Changes in CLU expression during chronic alcohol consumption have not been reported.
Not nCLU, but sCLU, has been suggested as a biomarker of neurodegenerative diseases, i.e., AD and PD, in the plasma or cerebrospinal fluid, respectively [40, 41]. In particular, CLU variants have been associated with AD in genome-wide association studies [54, 55], and plasma CLU levels reflect AD severity [40]. Therefore, CLU has been suggested as an AD risk gene. Intriguingly, sCLU, a pro-survival isoform, appears to fail to prevent the progression of neurodegeneration in AD, although it is upregulated in parallel to disease progression. In this context, sCLU might play a role as a chaperone in a similar manner to small heat shock proteins [56]. Expression and any contribution of nCLU need to be elucidated in neurodegenerative diseases as well. As nCLU plays a proapoptotic role, it is likely that nCLU expression could worsen neurodegeneration.
Concluding Remarks
In conclusion, we have discovered the mechanism of the nCLU pro-apoptotic function in vitro and in vivo, which shows that nCLU interacts with Bcl-XL though a putative BH3 domain. Notably, all BH3-only proteins interact with and regulate the core Bcl-2 family proteins to promote apoptosis, and multiple BH3-only proteins are usually involved in cell death in one cell type [30]. Thus, it would be interesting to know whether nCLU and other BH3-only proteins are cooperative in the cell death process.
sCLU upregulation in AD is already suggested as a useful diagnostic and prognostic biomarker. nCLU expression and upregulation could be used as a biomarker of cell death and brain damage as well. More importantly, cell death could be enhanced or reduced by regulating nCLU expression. For example, inhibiting nCLU may reduce brain damage by seizure or ethanol and upregulating nCLU may reduce tumors. However, CLU has two isoforms and multiple binding proteins besides Bcl-XL. Although previous results imply that regulating CLU isoforms could change cell fate, the biological roles of CLU and the mechanism of cell death/survival regulated by CLU in vivo are thought to be more complex, depending on many factors, such as the expression/induction of each isoform, subcellular location of the protein, various stimuli, and the microenvironment. Thus, there is an urgent need to develop isoform-specific antibodies and siRNAs for further clarification of the biological roles of CLU in various cell types and conditions.

 XML Download
XML Download